

Source: MDPI
1. Introduction
Scientific reports usually begin with a brief rationale which explains why the given topic was undertaken—studied/reviewed/thought of. In the case of this work, we strongly feel that in July 2020, COVID-19, the cause of severe respiratory failure and over 673,169 deaths worldwide, and at the same time, the origin of an upcoming global economic crisis, does not need to be introduced to anyone.
To the best of our knowledge, no specific and effective pharmacological treatment has yet been established, although more than 2834 clinical studies have already been registered (as of 30 July 2020).The core idea of this work was initially based on a link between two papers—one, which states that Zn(II) is a potent inhibitor of coronavirus RNA-based polymerase [1] and another, which shows that chloroquine, a common antimalarial drug, also used against several autoimmune diseases, is a zinc ionophore [2].
This core idea is sustained by: (i) the results of the first independent clinical trials which show the clinical benefit of chloroquine (CQ) and hydroxychloroquine (HCQ) treatment in COVID-19 infected patients [3,4], and further built on: (ii) numerous reports on the anticoronaviral and antiviral properties of CQ and HCQ, (iii) antiviral properties of Zn(II) and Zn(II) ionophores, and (iv) ongoing clinical trials which include Zn(II) in CQ/HCQ treatments.
Could the mode of action, or at least one of the possible modes of action of CQ/HCQ be as simple as being an ionophore, thus bringing more Zn(II), a potent RNA-based polymerase inhibitor, into the cell? At this place, it would be honest to add, that this question remains a rhetorical one also at the end of this paper—clinical studies that aim to answer it are still ongoing. Zinc(II) could turn out to be the éminence grise, but it could also turn out to be one of the pieces of a really big puzzle.
We do not provide the reader with a definite answer, but elaborate more on this idea, shedding light on Zn(II) as a possible RNA-dependent RNA polymerase inhibitor and on the possible anticoronaviral and antiviral mode of action of zinc(II) ionophores. We show the first success and the (impressive) amount of interest in chloroquine and hydroxychloroquine in COVID-19 clinical trials and also briefly summarize the massive amount of ongoing COVID-19 clinical trials on non-Zn(II) ionophore treatments. It is important to add that the latter task is not a trivial one, since the amount of COVID-19 clinical trials increases at an incredible rate; while many others are already in progress, not yet registered, but already approved by the corresponding bioethics committees.
The situation with the use of HQ itself as a COVID-19 drug is ‘very dynamic’, and even as this article was being written, several articles came out on the results of clinical trials, often ambiguous in their conclusions, ranging from extremely positive in a study involving 550 Chinese patients [5] to no benefit in a study on 181 French patients [6]. Such “dynamic” and “hot” topics are very likely to attract cases of scientific misconduct—this was the case of a famous Lancet paper by Mehra et al. [7], published on May 22nd, discussing a huge amount of medical data (over 96,000 patients from 671 hospitals) which, according to the authors, showed that chloroquine/hydroxychloroquine therapy was ineffective and, what is more distressing, increased the risk of ventricular arrhythmias and in-hospital deaths with COVID-19.
After that, the situation becomes “dynamic”: based on this publication, on May 25th, the WHO decides to withdraw some of the clinical tests with HQ/HCQ [8]. On June 3rd, Lancet publishes an Expression of Concern [9], which mentions the presence of “serious scientific questions” and errors in the article, and an audit is commissioned by three of the four authors not connected to Surgisphere—the company responsible for the data collection and analysis. Two days later, the same three of the four authors of the mentioned article ask the editor to retract the paper.
According to the authors’ announcement, after having been rejected access to the Surgisphere patients’ databases to the independent auditors, they could not vouch for the veracity of the data any longer [10]. At the same time, a similar situation happens to an New England Journal of Medicine paper on the safety concern of chloroquine [11], which relied on data from the same company which took part in the Lancet study.At the time this article was being written, several excellent preprints on the combination of CQ/HCQ came out: the first one by Scholz and Derwand, who also suggest that CQ and HCQ can inhibit SARS-CoV-2 replication by increasing pH in intracellular vesicles and by targeting extracellular zinc(II) to lysosomes, where Zn(II) then acts as an RdRp inhibitor [12].
They also point out that zinc(II) deficiency often occurs in those patients, who are at higher risk of a severe course of COVID-19—those with cardiovascular disease, chronic pulmonary disease, or diabetes. Another work by Scholz et al.
[13] describes outcomes of the treatment of COVID-19 patients after early treatment with zinc(II), low dose hydroxychloroquine, and azithromycin (the triple therapy) dependent on risk stratification. The study, which includes a treatment group of 141 COVID-19 patients (median age 58 years) treated with the triple therapy for 5 days, shows that the triple therapy treatment of COVID-19 should be included as early as possible after symptom onset. The zinc(II) and hydroxychloroquine treated group was associated with significantly less hospitalizations (84% less than in the untreated control group) and 5 times less all-cause deaths (one patient (0.7%) died in the treatment group versus 13 patients (3.5%) in the untreated group). The study further underlines the importance of the early onset of Zn(II) treatment, which makes perfect sense if we consider Zn(II) as an RNA-dependent RNA polymerase inhibitor (as discussed in Section 2).Another preprint shows the first in vivo evidence that zinc sulfate in combination with hydroxychloroquine may play a role in therapeutic management for COVID-19—administration of zinc sulfate together with HQ to 411 patients resulted in decreased mortality, admission to the ICU, and the need for ventilation [14].
This additionally encouraged us to look for more FDA-approved Zn(II) ionophores (or even those which are present in food) with antiviral, but also anticancer and antibacterial properties—because, if the mode of action of chloroquine and hydroxychloroquine really is based on their Zn(II) ionophoric activity, why not focus on much safer, clinically used, or naturally occurring zinc(II) ionophores with less side effects? In Section 5, Section 6 and Section 7, we summarize the current knowledge on FDA–approved Zn(II) ionophores with antiviral, anticancer and/or antibacterial activity. Could they be of any use in the anti-COVID fight?
2. Zn(II) as an RNA-Dependent RNA Polymerase Inhibitor
RNA-dependent RNA polymerase (RdRp) is the core enzyme in the replicative cycle of all positive-strand RNA (+RNA) viruses, which catalyzes the replication of RNA from a RNA template in a two-step mechanism [15,16,17]: (i) the initiation step of RNA synthesis begins at the 3′ end of the RNA template and the (ii) elongation phase, in which the nucleotidyl transfer is repeated with subsequent NTPs to generate the complementary RNA product.
Coronaviruses (order Nidovirales) are a family of +RNA viruses with the largest single-stranded RNA genomes (~30kB) known to date [18,19,20]. The unusual complex replication and transcription machinery of severe acute respiratory syndrome coronavirus (SARS-CoV) is composed of 16 nonstructural proteins (nsps), produced during co- and posttranslational proteolytic processing of two replicase polyproteins (pp1a and pp1ab). The canonical RdRp is localized in the C-terminal part of nonstructural protein 12 (nsp12) and possesses an architecture common to all viral polymerases.
Their activity is greatly stimulated by the nsp7 and nsp8 co-factors, and CoV RdRp which belongs to the class of primer-dependent polymerases. The overall architecture of the CoV nsp12-nsp7-nsp8 complex is well known and widely described in literature, and it was recently shown that SARS-CoV-2nsp12-nsp7-nsp8 complex has a conserved structure typical for SARS-CoV viruses (Figure 1a). The nsp7-nsp8 pair of SARS-CoV-2 shows a structure, similar to the SARS-CoV nsp7-nsp8 pair, with 13 additional amino acid residues only [21,22].
The nsp12 polymerase binds to the nsp7-nsp8 heterodimer with the second subunit of nsp8 occupying a distinct binding site. SARS-CoV-2 nsp12 contains a polymerase domain (a.a. 367–920) that adopts a cupped “right hand” like structure similar to other polymerases. The polymerase is composed of three subdomains: the fingers domain (a.a. 366–581 and 621–679), a palm domain (a.a. 582–620 and 680–815) and a thumb domain (a.a. 816–920) (Figure 1a).
SARS-CoV-2 nsp12 also contains a nidovirus-unique N-terminal extension domain (a.a. 60–249) that adopts a nidovirus RdRp-associated nucleotidyltransferase (NiRAN) architecture. These two domains are connected by an interface domain (a.a 250–365) (Figure 1a). The active site of SARS-CoV-2 RdRp has typically seven conserved motif regions (A–G) involved in template and nucleotide binding and catalysis.
The mechanism by which SARS-CoV-2 polymerases carry out de novo RNA initiation is similar to those described for SARS-CoV and for other RNA polymerases such as HCV and PV polymerase [22]. The incoming nucleoside triphosphate (NTP) enters the active site through a channel formed by a set of hydrophilic residues in motif F.
The RNA template is expected to enter the active site composed of motifs A and C through a groove clamped by motif F and G. Motif E and the thumb subdomain support the primer strand. The product-template hybrid exits the active site through the RNA exit tunnel at the front side of the polymerase [21,22].
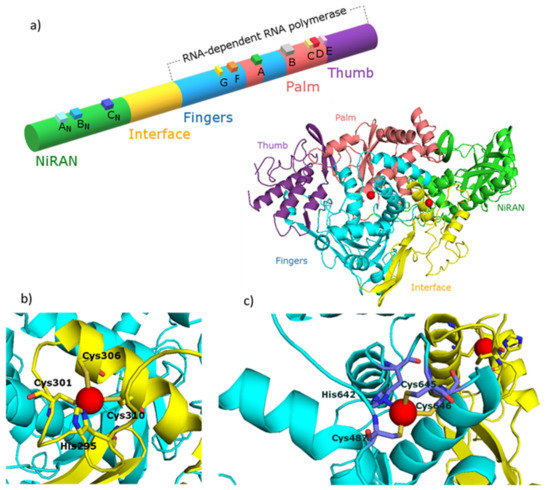
Figure 1. (a) Diagram and structure of the SARS-CoV nsp12 protein indicating protein domains (NiRAN, interface, fingers, thumb and palm), conserved motifs (AN, BN, CN, G, F, A, B, C, D) and zinc(II) binding sites (red spheres).
Enlarged zinc(II) binding sites of nsp12 protein placed in: (b) interface region and (c) fingers region. The figure was generated using PyMOL [41]. PDB entry: 7BTF [42]. Figures based on [21,22].Taking into account the essential role of the RdRp in the virus life cycle, as well as the effectiveness of polymerase inhibitors used in the treatment of other viral infections (HIV-1, HBV, HCV and herpes virus), the SARS-CoV-2 RdRp is widely recognized as one of the main, important and thus attractive targets for the rational design of anti-CoV drugs [23,24].
Zn(II) is naturally present in the structure of 2019-nCOV’s RdRp (Figure 1), and its elevated amounts clearly inhibit the course of infection. There is a growing amount of evidence that one of the promising candidates to have direct inhibitory effect on the replicative cycle of SARS-CoV-2 might be Zn(II).
Zn(II) is a generally known immune system enhancer as well as viral inhibitor—numerous studies have shown that intracellular Zn(II) levels have an effect on the life cycle of various RNA viruses that infect the human respiratory tract, including influenza [25], respiratory syncytial virus [26] and several picornaviruses [27,28,29,30,31,32]. Zinc(II) also inhibits the replication of DNA-containing viruses: human immunodeficiency virus-1 (HIV-1) [33], vaccinia virus [34], and polioviruses [35] in vitro.
Several potential mechanisms have been proposed for the observed zinc(II)-mediated inhibition of RNA viruses replication in vitro [29,36,37]; for some RNA viruses, the disruption of replication has been attributed to the interference with viral protein processing [32], while for other, the direct inhibition of RdRp activity has been suggested [1,38,39,40].Studies on the effectiveness of heavy metal chelating agents for their inhibitory effect on the influenza RdRp, reveal both surprising and promising results. Now, one could expect that chelating Zn(II) could, at least theoretically, inhibit the proper folding of RdRp; instead, the contrary was observed: when Zn(II) ions were added to the influenza RNA polymerase enzyme complex, they showed inhibitory activity, but the inhibition effect of the addition of Zn(II)-chelator complexes was greater than the effect of the chelating agent itself.
It was suggested that Zn(II)-chelator complexes dissociate in solution, after which Zn(II) is absorbed by proteins, including polymerase polypeptide(s), and thus exert a separate inhibitory effect on RdRp [25,38]. An inhibitory effect of Zn(II) ions with an IC50 (inhibitory concentration: median value of inhibitor, where 50% of biological and biochemical function of organism inhibition is observed) of about 60 mM on HCV RdRp activity was demonstrated.
The authors postulated two possible mechanisms of the observed scenario: (i) Zn(II) ions compete with Mg(II) and Mn(II) ions for binding to the carboxylate of three aspartic acids from A and C motifs, or (ii) zinc(II) ions bind to another site, different from the catalytic one, and form an allosteric obstacle caused directly by a nucleophilic attack on the α-phosphate of the 3′-hydroxyl group of the primer [39].
Another work by Hung et al. showed that Zn(II) inhibits HRV-16 3D polymerase with an IC50 as low as 0.6 mM, but the authors did not believe in a possible displacement of the catalytically essential Mg(II) ion due to the significantly lower concentration of Zn(II) compared to Mg(II) [40]. At this point it is important to underline that the discussed mechanisms are still hypotheses, and the exact mechanism has not yet been proved in detail.
The inhibitory effect of Zn(II) ions on RdRp was also observed for two major pathogens of humans and livestock, severe acute respiratory syndrome coronavirus and the arteriviruses equine arteritis virus (EAV), respectively [1]. Zn(II) concentrations as low as 2 µM appear to be a specific inhibitor of the initiation and elongation phases of RNA synthesis.
The authors suggested that Zn(II) does not compete with Mg(II) and binds to another Zn(II)-specific binding site [1]. In 2019 Kirchdoerfer et al. confirmed, for SARS-CoV, two metal binding-sites to which they have assigned Zn(II) binding. These binding sites are highly conserved across the CoV family [21].
As previously predicted for the SARS-CoV, also SARS-CoV-2 nsp12 contains two metal-binding sites to which Zn(II) ions are bound. The Zn(II) ions in both viruses are present in the same location: the first binding site is in the interface domain (His295, Cys301, Cys306 and Cys310) and the second is in the fingers domain (Cys487, His642, Cys645 and Cys646) (Figure 1b and c, respectively).
Both of these metal-binding sites are away from known active sites as well as protein–protein and protein–RNA interactions. Therefore, it is thought that Zn(II) ions play a structural role in the folded protein rather than are directly involved in enzymatic activity [21,22]. An in-depth structural analysis and subsequent mutational studies targeting aforementioned metal-binding sites are required to provide further insight into and a structural basis for the Zn(II)-induced inhibitory effects on RdRp activity.
What is clear so far is the fact that the combination of Zn(II) ions and Zn(II)-ionophores inhibits the nidovirus replication in cell culture. If the Zn(II) ions can inhibit coronavirus replication, then theoretically, there is just one challenge to overcome–to increase the concentration of Zn(II) in virus infected cells.
3. Anticoronaviral Properties of Chloroquine and Hydroxychloroquine
3.1. Zinc(II) Ionophores and Other Possible Modes of Action
In the context of searching for substances that help to transport Zn(II) ions from the external environment into the cell, all eyes are set on chloroquine and its less toxic analogue, hydroxychloroquine [4,43,44].
Chloroquine, discovered more than 80 years ago, has been used in clinical practice for over 70 years and its patent has expired (in other words “it is cheap”) and has a reasonably safe clinical safety profile, with the side effects of its intake being severe cardiac arrhythmia/heart failure, hepatic disorders, headache, dizziness and gastrointestinal disorders. It also has to be taken into account that chloroquine interacts with many other drugs (azithromycin, cyclosporine, amiodarone, penicillamine, digoxin, cimetidine) and its intake must be strictly monitored in hospitals [45].
Through the years, it has been used to treat malaria and several other diseases, such as rheumatoid arthritis or systemic lupus erythematosis [46]; although some hypothesis exist, its modes of action against these diseases (especially the autoimmune ones) have not been yet sufficiently understood [47,48,49].
It may be correlated with the inhibition effect of phospholipase A2, interaction with lysosomes, inhibition of phagocytosis, inhibition of peroxide synthesis, increase of intracellular pH and reducted stimulation of CD4 lymphocytes. Chloroquine can also interact with porphyrins, acting as an inhibitor of the biocrystallization of hemozoin (a disposal product formed from the digestion of haemoglobin, non-toxic for parasites) [47,48,49].
Now (30.07.2020), with 329 ongoing, promising CQ and/or HCQ anti COVID-19 clinical trials, it is crucial to find out the precise mode of their anti-COVID-19 action in order to be able to design more suitable treatments with less side effects.Studies performed in 2014 by Xue et al. [2] (we are now referring to the paper which started the avalanche of Zn(II)-ionophore-anti COVID-19 discussions) brought new insight into chloroquine’s activity, reporting significantly elevated intracellular Zn(II) levels when both Zn(II) and chloroquine were added to the cell culture medium, which indicates that chloroquine may act as a Zn(II) ionophore that targets Zn(II) ions into the cell and/or into lysosomes.Chloroquine and hydroxychloroquine have also been reported to be antiviral agents in papers, where the use of Zn(II) is not mentioned [50,51].
So far, the effectiveness of chloroquine or hydroxychloroquine has been tested against five of seven identified human coronaviruses. They are members of two viral genus: alphacoronaviruses (Human coronavirus HCoV-229E) and betacoronaviruses (SARS-CoV, SARS-CoV-2, MERS-CoV and HCoV-OC43). A simplified classification of the mentioned viruses is presented in Figure 2 [52].

Figure 2. Classification of chosen coronaviruses. Species described in this section were marked in red.Chloroquine can inhibit SARS-CoV infection at micromolar (IC50 = 8.8 µM) concentrations. Interestingly, this IC50 is similar to the chloroquine concentration in plasma during malaria treatment [53].
In the suggested mechanism of action, chloroquine (a weak base) causes an increase of endosomal and lysosomal pH, which inactivates the host’s pH-dependent enzymes necessary for viral entrance inside the host cell [51,54,55]. To enter, SARS-CoV interacts with angiotensin-converting enzyme 2 (ACE2) receptor by the spike (S) proteins.
Two functional domains of these proteins can be distinguished—S1, responsible for receptor binding and S2—for viral and cellular membrane fusion with specified fusion peptide region, interacting with lipid bilayer [56,57].
After virion uptake, the S protein is specifically cleaved to expose the fusion peptide region. Enhancing the lysosomal pH, chloroquine inactivates pH-dependent lysosomal proteases like cathepsin L necessary for S-protein cleavage, finally preventing the membrane fusion and viral genome release [56,58].
Chloroquine and hydroxychloroquine reduce the glycosylation of the ACE2 receptor and the viral penetration into the cell, due to the alcalisation of endocytic compartments—when not glycosylated, they will have lower affinity towards the viral spike proteins, which probably impedes the virus—receptor binding, resulting in the inhibition of the infection development [55].
Moreover, in in vitro experiments, chloroquine, administered before exposure to the virus, significantly reduces the development of infection, which indicates not only therapeutic but also prophylactic activity. CQ and HCQ caused an immunomodulating effect and it is assumed that the cytokine storm of SARS-CoV-2-infected patients suffering from COVID-19 will be inhibited [55].
Due to the similarity between the SARS-CoV-2 and SARS-CoV viruses, chloroquine’s mechanism of action against them will most likely be similar. In the case of SARS-CoV-2, in vitro assays show that chloroquine and hydroxychloroquine’s antiviral activity is low (it reaches micromolar concentrations) when administered both before and after viral infection (entry and post-entry stage) [43,59].
Furthermore, both agents inhibit the viral transfer from endosomes to lysosomes, which appears necessary for the release of viral genome [43]. Moreover, CQ and HCQ have anti-inflammatory properties, which may alleviate the inflammatory response in in vivo experiments [43].
Clinical trials done in China have shown that chloroquine is effective against SARS-CoV-2-induced infection, alleviating and shortening the disease course with no severe unfavorable effects [3]. Also clinical trials carried out in France on 20 patients (200 mg hydroxychloroquine sulfate, three times per day during 10 days) have shown that 70% of them were virologically cured 6 days after drug inclusion, compared to 12.5% in the control group [4].
Right now, there are 329 ongoing anti-COVID-19 clinical trials (85 on chloroquine and 244 on hydroxychloroquine)–they are commented on in the next section [60].In vitro studies of chloroquine and hydroxychloroquine on other coronaviruses showed that chloroquine was effective against MERS-CoV at an early step of infection (statistically calculated effective concentration that induces a specific effect in 50% of experimental population under direct/specific conditions, EC50 = 3.0 µM) [61].
Human coronaviruses HCoV-OC43, together with HCoV-NL63, HCoV-229E and HCoV-HKU1 induce approximately 15% of common cold infections all over the world [62,63]. In vitro tests show that HCoV-OC43 is also susceptible to chloroquine (EC50 = 0.306 µM).
In vivo tests, in the murine model, confirmed chloroquine’s effectiveness against HCoV-OC43, when administered at the initial stage of infection [64]. Replication of another human coronavirus, HCoV-229E, was significantly decreased by chloroquine at 10 and 25 µM concentrations. In this study, chloroquine was suspected of inhibiting the activation of p38 mitogen-activated protein kinases (p38 MAPKs) in infected cells [65].
Many viruses are able to use MAPKs to adapt cellular processes to their own purposes, thus inhibiting the activation of p38 MAPK may reduce viral replication [65,66].
In addition to human viruses, chloroquine antiviral activity was also tested against animal coronaviruses, i.e., feline infectious peritonitis virus (FIPV), also known as feline coronavirus (FCoV), which belongs to the alphacoronaviruses and is responsible for inducing the feline infectious peritonitis (FIP)—a fatal disease in domestic and wild cat species [67,68].
Chloroquine decreased viral replication, which was confirmed by both in vitro and in vivo experiments. Due to the anti-inflammatory effect, chloroquine was suggested as a potential therapeutic agent in FIP treatment [68].
3.2. Ongoing Clinical Trials
To date (30th July 2020) there are 2834 clinical studies concerning COVID-19 [60]. Chloroquine and hydroxychloroquine (alone or combined with other interventions such as: other drugs, dietary supplements, biological agents, medical devices and others) are considered as treatment in 329 studies (85 on chloroquine and 244 on hydroxychloroquine (where also some CQ ones are included).
Chloroquine/chloroquine phosphate/chloroquine diphosphate and/or hydroxychloroquine are most frequently combined with: (i) anti-infective agents which have been already approved and used as:
Antibiotics (azithromycin, ceftriaxone, piperacillin-tazobactam, ceftaroline, amoxicillin-clavulanate);
Other antibacterial drugs (moxifloxacin, levofloxacin, carrimycin);
Drugs used to treat and prevent HIV/AIDS (darunavir/cobicistat, lopinavir/ritonavir and emtricitabine/tenofovirdisoproxil);
Other antiviral drugs (remdesivir, oseltamivir, interferon-β1a);
Receptor antagonists (losartan, anakinra, sarilumab) and others (inhibitor of Janus kinase—baricitinib; (ii) antiinflamatory and immunosuppressive agents (hydrocortisone, tocilizumab); (iii) respiratory system agents (expectorants, bromhexine);
(iv) anti-arrhythmia/antihypertensive agents (losartan);
(v) anticoagulants (gabexate); (vi) anti-allergic agents (ciclesonide);
(vii) antineoplastic agents (topoisomerase inhibitors, pembrolizumab, ofloxacin, moxifloxacin, levofloxacin, interferons);
(viii) fibrinolytic agents (defibrotide); (ix) renal agents (ofloxacin, levofloxacin);
(x) vasoconstrictor agents (giapreza, angiotensin II) and (xi) micronutrients (ascorbic acid, folic acid, ergocalciferols, hydroxocobalamin, iodine, vitamin D, zinc(II)).
Especially interesting (in particular from the point of view of this review) is the last group, where CQ/HCQ is considered as a treatment, together with zinc(II) and other micronutrients. Eighteen of all clinical studies assume that including zinc(II) as a dietary supplement may be essential for achieving effective treatment or prevention of COVID-19.
A brief overview of those studies is given in Table 1 below.
Table 1. Characteristics of clinical trials concerning zinc(II) as a treatment of COVID-19.
| Main Drug | Other Interventions | Clinical Trial Identifier | Duration | Location | Purpose | Estimated Enrollment | Inclusion Criteria: | Ref. |
|---|---|---|---|---|---|---|---|---|
| Hydroxychloroquine | Vitamin C, vitamin D, zinc(II) | NCT04335084 | 24 weeks | Ventura, CA, USA | prevention | 600 participants, medical workers who are exposed to COVID-19 | Patients >18 years old without COVID-19 symptoms; male or female that are considered to be high-risk individuals | [69] |
| Hydroxy-chloroquine (Plaquenil) (200 mg) | Vitamin C, vitamin D, zinc(II) | NCT04326725 | 16 weeks | Istanbul, Turkey | prevention | 80 participants, healthcare professionals and their first degree relatives | Patients aged between 20 and 90 years without COVID-19 symptoms; male or female | [70] |
| Hydroxy-chloroquine (Plaquenil) | Azithromycin (Zithromax), vitamins C and D Zinc(II) | NCT04482686 | 24 weeks | Ventura, CA, USA | treatment | 600 participants, patients with COVID-19 infection | Patients aged between 18 and 75 years; healthy male or female with positive test for COVID-19; diabetic and obese (BMI > 30) patients will be included in the trial but randomization will be stratified | [71] |
| – | Oral nutrition supplement (ONS): protein (14800 mg), fat (22200 mg), carbohydrate (25000 mg), EPA (1100 mg), DHA (450 mg), GLA (950 mg), vitamins A (2840 IU), C (205 mg), and E (75 IU), selenium (0.018 mg), zinc(II) (5.7 mg) | NCT04323228 | 12 weeks | King Saud University Saudi Arabia | supportive care | 30 participants patients with COVID-19 infection | Patients aged between 18 and 65 years; male or female in stable condition (i.e., not requiringICU admission) | [72] |
| – | Vitamin C (8000 mg) Zinc(II) Gluconate (50 mg) | NCT04342728 | 4 weeks | Cleveland, OH, USA | supportive care | 520 patients diagnosed with COVID-19 | Male or female patients >18 years old with COVID-19 diagnosed in an outpatient setting with any symptom (fever or chills, shortness of breath or difficulty breathing, cough, fatigue, muscle/body aches, headache, new loss of taste, new loss of smell, congestion or runny nose, nausea, vomiting, diarrhea | [73] |
| Hydroxy-chloroquine (400 mg, 200 mg)/azithromycin (500 mg, 250 mg) hydroxy-chloroquine (400 mg, 200 mg)/doxycycline (200 mg) | Zinc(II) (220 mg) | NCT04370782 | 8 weeks | Roslyn, NY, USA | treatment | 750 participants with COVID-19 infection | Patients 30 years and older; male or female with high initial clinical suspicion by physician based on signs and symptoms (fever, cough, myalgias, fatigue, shortness of breath) | [74] |
| Hydroxy-chloroquine (400 mg) | Zinc(II) (15 mg) | NCT04377646 | 8 weeks | Tunis, Tunisia | prevention | 660 participants military health professionals exposed to SARS-CoV-2 | Patients aged between 18 and 65 years; male or female without COVID-19 symptoms and with negative SARS-CoV-2 diagnostic test; no self-medication with chloroquine, hydroxychloroquine or antivirals | [75] |
| – | Vitamin D3 (2000 IU) Zinc(II) (30 mg) | NCT04351490 | 8 weeks | Lille, France | treatment | 3140 participants, 60 years and older patients infected with COVID-19 | Patients 60 years and older; male or female; institutionalized | [76] |
| Hydroxy-chloroquine, azithromycin, favipiravir (1800 mg, 800 mg) | Zinc(II) | NCT04373733 | 4 weeks | London, UK | treatment | 450 participants infected with COVID-19 | Male or female patients > 18 years old with suspected or confirmed COVID-19 infection (fever ≥37.8 °C and at least one of the following respiratory symptoms, which must be of acute onset: persistent cough, hoarseness, nasal discharge or congestion, shortness of breath, sore throat, wheezing or sneezing) | [77] |
| Chloroquine | Zinc(II) | NCT04447534 | 2 weeks | Tanta, Egypt | treatment | 200 participants infected with COVID-19 | Patients 18 years and older with positive COVID-19 (inclusion/exclusion criteria not very precise; expected treatment outcome difficult to predict) | [78] |
| Hydroxy-chloroquine (400 mg, 200 mg), azithromycin (500 mg, 250 mg) | Vitamin C (400 mg/kg/day, 200 mg/kg/day Zinc(II) (30 mg) Vitamin D3 (5000 IU) Vitamin B12 (0.5 mg) | NCT04395768 | 18 months | Melbourne Australia | treatment | 200 participants infected with COVID-19 | Patients who were > 18 years old; male or female with active COVID-19 infection | [79] |
| Hydroxy-chloroquine (800 mg, 400 mg) | Zinc(II) (66 mg) | NCT04384458 | 7 weeks | Fortaleza, Ceará, Brazil | prevention | 400 healthcare workers exposed to SARSCoV-2 | Patients aged between 18 and 70 years; male or female which works in a healthcare facility delivering direct care to patients with either proven or suspected COVID-19 | [80] |
| Hydroxy-chloroquine (400 mg; 200 mg) | Ivermectin (12 mg) Povidone-Iodine Zinc(II) (80 mg) Vitamin C (500 mg) | NCT04446104 | 6 weeks | Singapore | prevention | 5000 participants, migrant workers at High-risk of COVID-19 | Patients aged between 21 and 60 years; male with weight more than 40 kg | [81] |
| Nitazoxanide, ribavirin, ivermectin | Zinc(II) | NCT04392427 | 2 years | MansouraEgypt | treatment | 100 participants infected with COVID-19 | Patients 12 years and older; male or female with the COVID-19 PCR positive test and referred to the quarantine; patient without any comorbidities and no sensitivity or contraindication to the three drugs | [82] |
| Hydroxy-chloroquine (200 mg) | Azithromycin (500 mg, 250 mg) Zinc(II) (220 mg) | NCT04412746 | 3 months | Algeria | treatment | 100 participants with diabetes and infected with COVID-19 | Patients 16 years and older; male or female; hospitalized patients with diabetes known before the admission; new onset diabetes discovered at admission | [83] |
| Quercetin (500 mg) | Bromelain (500 mg) Zinc(II) (50 mg) Vitamin C (1000 mg) | NCT04468139 | 4 weeks | Saudi Arabia | treatment | 60 participants infected with COVID-19 | Male or female patients aged ≥ 18 years with moderate to severe disease (inclusion/exclusion criteria not very precise) | [84] |
| Ivermectin (0.2 mg/kg) | Nigella Sativa (80 mg/kg) Zinc(II) (20 mg) | NCT04472585 | 4 weeks | Punjab, Pakistan | treatment | 40 participants infected with COVID-19 | Male of female patients > 18 years old with a positive SARS-CoV-2 PCR test; patients with mild to moderate disease | [85] |
| Hydroxy-chloroquine | Vitamin C (2000 mg) Vitamin D (Alfacalcidiol 0.001 mg) Zinc(II) (50 mg) Paracetamol | NCT04491994 | 3 weeks | Punjab, Pakistan | treatment | 500 participants infected with COVID-19 | Patients aged between 18 and 80 years; male or female, with mild disease | [86] |
Among the 2834 clinical studies concerning COVID-19 (30.07.2020), 743 of them relate to drug interventions (among them are the 329 chloroquine and hydroxychloroquine ones described above) [60]. Others include: biological studies (for example natural killer (NK) cells, recombinant vaccines, mesenchymal stem cells (MSCs) or convalescent anti-SARS-CoV-2 plasma, diagnostic tests (for example breath test, serological tests, lung ultrasound, new QIAstat-Dx fully automatic multiple PCR detection platform, SARS-CoV-2 IgG Antibody Testing Kit), devices (e.g., BIOVITALS, biosensors, SensiumVitals wearable sensor), different questionnaires and others (e.g., SPIN-CHAT Program or telerehabilitation).
Data from ClinicalTrials.gov [60] show that the largest number of drugs tested are included in the category of anti-infective agents (over 16%), but a significant part are also antihypertensive agents (8.3%), antineoplastic agents (7.0%), central nervous system depressants (5.1%), vasodilator agents (4.4%), anti-inflammatory agents (4.3%), analgesics (4.0%), antirheumatic agents (3.8%), micronutrients (3.6%), gastrointestinal agents (3.4%), anticoagulants (3.1%).
The remaining, approximately 40% are drugs belonging to a variety of categories, including among others: respiratory system agents, channel blockers, lipid regulating agents, cardiotonic agents, dermatologic agents, neuroprotective agents and others (shown in Figure 3).
Figure 3. Potential drugs involved in clinical trials against COVID-19 classified in individual categories [60].As stated above, hydroxychloroquine, chloroquine and chloroquine diphosphate are some of the most frequently studied. Other drugs, which are not dependent on zinc(II) ions, and which appear in numerous clinical studies are comprehensively listed in the Supplementary Materials (Table S1).
Non-CQ/HCQ and non-Zn(II) involving in vitro studies are now mainly focused on:(i)S protein-ACE2-mediated system and thus viral entry [87,88],(ii)compounds targeting SARS-CoV main protease [89,90],(iii)agents targeting papain-like protease 2 (PLP2) [91],(iv)agents targeting SARS-CoV RdRp system [92,93], and(v)agents targeting SARS-CoV helicase [94].
4. Antiviral Properties of Chloroquine and Hydroxychloroquine
Chloroquine and its derivatives have not only an enormous impact on the treatment of coronaviruses [53,55,59,64] (see the previous section), but are also effective against several others viruses in vitro, including: (i) human immunodeficiency virus (HIV) [95,96,97,98,99,100,101], (ii) hepatitis A (HAV) [102], B (HBV) [103], C (HCV) [104] viruses and autoimmune hepatitis [105], (iii) poliovirus [106], (iv) influenza A [107,108], B [109] and H5N1 [110] viruses, (v) Zika virus (ZIKV) [111,112,113], (vi) Ebola virus (EBOV) [114], (vii) Crimean–Congo hemorrhagic fever virus (CCHFV) [115], (viii) rabies virus [116], (ix) Chikungunya virus (CHIKV) [117,118], (x) Dengue virus (DENV) [119,120,121], (xi) Hendra, Nipah viruses [122], and (xii) herpes simplex virus (HSV) [123].
In this section, we briefly discuss the possible “non-Zn(II)-based” modes of action (if known) of chloroquine and hydroxychloroquine.Among the above mentioned examples, the most comprehensively described antiviral effects of chloroquine and hydroxychloroquine were those against HIV replication, what was tested both in clinical trials [124,125] as well as in in vitro studies.
Cell line models [100,126], peripheral blood lymphocytes and monocytes [101], were used to examine chloroquine/hydroxychloroquine inhibition effect on HIV. The mechanism of anti-HIV effect of chloroquine may be associated with several different pathways: (i) inhibition of HIV-1 integrase, (ii) inhibition of Tat(viral early protein)-mediated transactivation, which leads to the decrease of transcription initiation, (iii) reduction of iron concentrations within the cell and thus affecting the reverse transcription [96,127,128,129], (iv) inhibition of HIV-1 replication, which is affected by the post-transcriptional maturation of glycoprotein gp120 [130], fundamental for viral infectivity.
Moreover, Savarino et al. showed that at clinically achievable concentrations, chloroquine reveals a broad-spectrum of anti-HIV-1 and HIV-2 activity, which results from the decrease of the incorporation of glucosamine in the envelope glycoproteins from both HIV-1- and HIV-2-infected cells [100]. This glycosylation process may become a new target for antiretroviral therapies.
Chloroquine was also examined in the in vitro treatment against Dengue virus type 2 (DENV-2), which leads to dengue fever [120], showing: (i) a significant reduction of virus production in infected cells, contrary to untreated cells, (ii) reduction of DENV-2 replication in cells, and (iii) reduction of proinflammatory cytokine expression [120]. Two clinical trials evaluated the efficacy of chloroquine treatment in patients infected with Dengue virus [131,132], showing that chloroquine does not reduce the duration of viraemia and antigenaemia in dengue patients; however, it does reduce fever clearance time and the incidence of dengue haemorrhagic fever [131].
The second clinical trial revealed the reduction of pain symptoms in DENV-infected patients and an improvement of daily fine fettle during treatment, but, on the other hand, did not alter the duration of the disease or the intensity and days of fever [132].
Other antiviral properties of chloroquine are mostly based on in vitro studies and have not been verified in clinical trials or have not revealed the satisfactory therapeutic effect. In hepatitis C virus infection, chloroquine inhibits the intracellular replication of an HCV replicon in a dose-dependent manner and could be used in hopeful combination therapy for HCV–HIV co-infected patients [104].
In case of influenza viruses, chloroquine have shown: (i) inhibitory effect against the replication of human influenza A virus H1N1 and H3N2 (in vitro but not in vivo) [107,108], (ii) the inhibition of influenza B infection by acting on a replicative step preceding primary transcription of the B virus genome by the addition of the drug at the time of infection (the drug had no effect on virus replication when added after 15 min after infection) [109], (iii) the inhibition of autophagy in mouse lung induced by avian influenza AH5N1 virus, while the viral proinflammatory cytokines were not significantly affected [110].
Chloroquine also inhibits Chikungunya alphavirus (CHIKV) infection in vitro, blocking the production of proinflammatory cytokines and the proliferation of monocytes, macrophages, and lymphocytes [117]. The effect of chloroquine on polioviruses is more modest—it can only redirect the processing of input virions without causing viral uncoating [106].
Chloroquine also has an antiviral activity against Zika virus (ZIKV) in different cell modes: Vero cells, human brain microvascular endothelial cells, human neural stem cells, and mouse neurospheres [111]. These studies prove both the reduction of the number of ZIKV-infected cells in vitro and the inhibition of virus production and cell death without a cytotoxic effect [111].
Promising in vitro results were also obtained for the Crimean-Congo hemorrhagic virus (CCHFV) in the presence of two FDA approved molecules: chloroquine and chlorpromazine [115]. The two drugs inhibited CCHFV virus replication even when given up to 6 -24 h after infection.Chloroquine has also been tested on animal models. Daily treatment of chloroquine to Ebola virus (EBOV) infected mice caused a significant increase of their survival rate (approximately up to 80%–90%) [133].
On the other hand, Dowall et al. [114] have shown that the replication of EBOV virus is indeed inhibited by chloroquine in vitro, but they did not observe any therapeutic effects on EBOV-infected guinea pigs in vivo. Similar, not effective chloroquine treatment in vivo was observed for Nipah and Hendra viruses (paramyxoviruses that cause severe disease such as encephalitis in animals and humans), in golden hamster models [122].
5. Other Antiviral Zn(II) Ionophores
Chloroquine and hydroxychloroquine are not the only antiviral zincophores – there are numerous other examples where inhibition of viral replication is observed (mostly in vitro). The replication of various viruses, including: (i) respiratory syndrome coronavirus, SARS-CoV [1], (ii) arteriviruses (equine arteritis virus, EAV [1], (iii) respiratory syncytial virus [26], (iv) porcine reproductive and respiratory syndrome virus, PRRSV [134] (v) picornavirus (poliovirus, Human rhinovirus, Foot-and-mouth disease virus (FMDV)) [27,30,31,32],(vi) herpes simplex virus (HSV) [135] and (vii) influenza virus [25], is inhibited by the simultaneous combination of two approaches: (i) high zinc ion concentration and (ii) the addition of compounds that induce cellular import of Zn(II), such as hinokitol (HK), pyrrolidine dithiocarbamate (PDTC) or pyrithione, (PT) (Figure 4).
The molecular mechanism of viral inhibition is most likely based on the rapid import of extracellular Zn(II) ions into the cells by ionophores, after which the imported Zn(II) ions inhibit the RNA polymerase activity and thus, replication [1,32,134].
Figure 4. An overview of zinc(II) ionophores discussed in this work: chloroquine (CQ), hydroxychloroquine (HCQ), hinokitol (HK), dithiocarbamates (DTCs), pyrrolidine dithiocarbamate (PDTC), pyrithione (PT), 8-hydroxyquinoline (PBT2), clioquinol, quercetin and zincophorin.
5.1. Pyrithione
For herpes simplex virus (HSV), a more detailed mechanism of inhibition by pyrithione (PT, 1-hydroxypyridine-2-thionine, omadine, Figure 4) was described [135]: (i) PT inhibits HSV late gene expression (e.g., that of glycoprotein D) and viral progene production, in a Zn(II) concentration dependent manner; (ii) PT blocks the HSV immediate early gene expression, and (iii) PT administration may inhibit the cellular ubiquitin-proteasome system (UPS), leading to the interruption of IκB-α degradation and NF-κB activation thus causing the reduction of viral infection symptoms. Pyrithione also shows anticancer, antimicrobial and antifungal properties, which are described in the next sections.
5.2. Quercetin
Quercetin (3,3′,4′,5,7-pentahydroxyflavone, Figure 4), found in onions, many Chinese herbs, vegetables, fruits and wines [136], is another zinc(II) ionophore, which shows antiviral effects against following viruses: (i) influenza A virus [137,138], (ii) hepatitis B (HBV) [139] and C (HCV) [140,141] viruses, (iii) Ebola virus (EBOV) [142], (iv) Dengue virus (DENV) [143], (v) Japanese Encephalitis Virus (JEV) [144] and (vi) Zika virus (ZIKV) [145].
The development of an influenza infection is initiated by hemagglutinin (HA), an influenza virus envelope protein, which helps in viral entry into the cell [146]. Mature HA is a homotrimer, and each monomer is consisted of two disulfide-linked subunits, HA1 and HA2 [146]. The mechanism of influenza A virus inhibition by quercetin is strictly related with virus entry into the cell [138].
Quercetin interacts with influenza hemagglutinin protein (most likely with the HA2 subunit) and thus inhibits the formation of the virus-cell link (in vitro studies) [138].
In case of hepatitis viruses, quercetin is responsible for Hepatitis B surface antigen (HBsAg), Hepatitis B antigen (HBeAg) and viral DNA level reduction in vitro. Furthermore, treatment with quercetin also decreases HCV virions integrity, HCV NS3 protease effectiveness and viral transcription levels [139,140,141].
Quercetin has also shown an antiviral efficiency in combination with other compounds. Houttuynia cordata Thunb. (HCT) extract, administered together with quercetin, quercetrin and cinanserin mixture showed an inhibitory effect on mouse hepatitis virus (MHV) and Dengue virus type 2 (DENV-2)in vitro [143]. Baicalein and quercetin combination may decrease the amount of the copied RNA of Japanese encephalitis virus [144].
Quercetin derivatives also exhibit effective antiviral activity. Quercetin 3-β-O-D-glucoside was successfully tested against Ebola [142] and Zika [145] viruses, nevertheless, further studies should be performed to elucidate the mechanism of its antiviral action.
5.3. Dithiocarbamates
Dithiocarbamates (DTCs, Figure 4), less-known Zn(II) ionophores, exhibit a broad spectrum of antitumor activities [147,148], and may serve as a potential anti-HPV virus drug candidates [149]. Moreover, diethyldithiocarbamate (DDTC), a derivative of DTCs, was also successfully used in patients with HIV-1 infection to delay progression of AIDS in clinical trials [150]. Zincophorin (Figure 4), a polyoxygenated ionophoric antibiotic, first isolated from Streptomyces griseus in 1984 [151,152], is active against Gram-positive bacteria and Clostridium coelchii in vivo (more details are given in Section 7) [152,153]. Interestingly, its methyl ester, exhibits a potent inhibitory properties against influenza virus with decreased toxicity for the host cells [154,155].
6. Anticancer Properties of Zinc(II) Ionophores
Zinc ionophores have a possible role not only in antiviral therapy, but also show anticancer and antibacterial properties. In this section, we discuss the most commonly used and most promising anticancer ionophores, leaving the reader with an open question—could they also be of any use in anti-COVID-19 therapy?
Metal ion homeostasis is crucial for the survival of both normal and cancer cells. Therefore, metal-binding compounds have been considered as potential anticancer agents [156]. Zn(II) ions are involved in cellular signal transduction, DNA synthesis and gene transcription, and are also essential for cell proliferation and differentiation [157].
Increased intracellular zinc concentration stabilizes hypoxia-inducible factor-1 (HIF-1) and therefore has been associated with glycolysis, angiogenesis and apoptotic cell death [158]. For these reasons, Zn(II) homeostasis is targeted to develop novel therapeutic agents for cancer treatments [159].
Two different mechanisms of ionophore-based anticancer activity can be distinguished. Some compounds acts via metal ion chelation. The chelators may bind the metals extracellularly, deplete the metal ions from cancer cells or inhibit molecular pathways, without necessarily removing metal ions [160,161,162]. Other anticancer drugs increase intracellular metal concentrations [159]. Here, we focus on the second mode of action and discuss Zn(II) ionophores.
6.1. Pyrithione
Pyrithione is a well-known antimicrobial and antifungal agent [163]. Its anticancer activity was first reported in the 1980s. However, due to its low water solubility (2.5 g/L at 20 °C) and poor bioavailability it is not appropriate for parenteral administration and is used only as a topical agent [164]. Nevertheless, several water-soluble derivatives have been synthesized so far.
They increase the pool of free Zn(II) ions and exhibit an anti- proliferative activity in A549 human lung cancer cell lines [158]. The treatment to be effective, it requires Zn(II) supplementation. This approach may be particularly important for prostate cancer – in normal healthy tissues, relatively high levels of Zn(II) ions are found, but this pool decreases drastically upon malignant transformation [165].
6.2. Clioquinol
It was shown that also clioquinol (5-chloro-7-iodo-8-hydroxyquinoline, Figure 4) acts as an ionophore and induces apoptosis of human cancer cells [166]. It should be kept in mind that clioquinol, as a metal chelator, possesses antimicrobial activity and is considered as a potential therapeutic in Alzheimer’s disease [167]. Although clioquinol induces inhibition of superoxide dismutase-1 (SOD1), the addition of metals essential for the enzyme’s activity, (which were expected to reverse the activity of the—presumably—sequestering agent) does not diminish cell damage, but on the contrary, enhances cell apoptosis.
The obtained results do not prove its metal chelation mode of action hypothesis, and strongly suggest that the addition of clioquinol increases the intracellular zinc pool, suggesting it acts as a zinc ionophore [166].
At this point it is worth to mention, that clioquinol has an interesting combination of properties, namely moderate metal affinity and intermediate pKa value. It allows metal binding outside the cell where a higher concentration of transition metal ions is observed (and also a higher pH), helping them to cross the cell membrane and then releasing metal ions where their concentration is lower (or a decreased pH is observed/competitive ligands are present).
It may be especially important in the treatment of cancer—within neoplastic cells, lower pH and increased concentration of glutathione, comparing to healthy cells, is always observed [168]. Therefore, clioquinol has an anticancer effect both in vitro and in vivo (e.g., on DU 145, a human prostate cancer line, and several other human cancer cell lines) [166], however, its mode of action is not really clear.
It has been shown that increased concentrations of free zinc(II) occurs in lysosomes, what may suggest that these organelles are primary targets of ionophores. Moreover, the addition of clioquinol and Zn(II) results in the degradation of BH3-interacting domain—death agonist (proapoptotic single domain BH3 proteins which act as apoptotic signal amplifiers or programmed cell death pathway activatiors), which plays a major role in lysosome-mediated apoptotic cell death. Clioquinol treatment results in the increase of free Zn(II) in lysosomes what leads to their disruption, thus apoptotic cell death of cancer cells is induced [169].
Schimmer and coworkers have recently demonstrated that clioquinol inhibits the proteasome through a copper-dependent mechanism and therefore inhibits growth of leukemia and myeloma cells [170]. It was also found that it inhibits NF-κB (nuclear factor κB) activity in ovarian cancer cells [171].
6.3. Dithiocarbamates
Another group of metal-binding drugs, which are capable to transport ions across cell membranes, are dithiocarbamates (Figure 4). They are widely used as insecticides, fungicides, therapeutic agents for alcoholism and metal intoxication [172], acting as a metal chelators and SOD1 inhibitors [173,174] or interacting with transcription factors like NF-κB and p53 [175,176,177].
Some dithiocarbamates, such as (the also antiviral and antibacterial) pyrrolidine dithiocarbamate (PDTC, Figure 4), are considered as a copper ionophores that recruit extracellular copper [178], what leads to the oxidation of endogenous thiols or conformational changes of p53 [176]. It was also shown that PDTC causes cerebral endothelial cell death. Transition metal ions, such as copper or zinc, may enhance the cytotoxic effect of PDTC [179]. It is worth noticing that the potency cytotoxic enhancement of zinc is probably greater than that of copper.
7. Antibacterial Zn(II) Ionophores
Ionophore antibiotics constitute a heterogeneous group of antimicrobials which are produced by microorganisms, mainly by various species of Streptomyces (Gram-positive spore-forming bacteria). Like other ionophores, they transport ions across cell membranes. It leads to the disruption of cell membrane permeability and results in antibacterial effects [180].
Ionophores are active only against Gram-positive bacteria, since Gram-negative bacteria have an outer membrane, acting as a permeability barrier [181].Widely used antibacterial group 1 and 2 ionophores such as monensin, salinomycin, narasin, lasalocid, maduramicin and laidlomycin transport alkali and/or alkaline-earth metal cations [180].
In all of the listed ionophores, metal ion binding is supported by intramolecular hydrogen bonds formation between the carboxylic and the hydroxyl groups. The hydrophilic cage is formed for the metal, which becomes surrounded by the hydrophobic part of the ionophore [182].Ionophores transferring cations belonging to group 1 and 2 of the periodic table are not the only ones exhibiting bactericidal activity. Significant interest has been raised to the antibacterial properties of transition metal complexes with ionophores [183].
Among them, Zn(II) ionophores seem to be those of potentially large application. An increase of the Zn(II) pool above several orders of magnitude above physiological level perturbs zinc(II) homeostasis and leads to a cytotoxic effect on prokaryotes at the concentration above 10−4 M [184,185]. It may also act as an antifungal agent, but in general, a higher ionophore concentration is needed to inhibit the growth of fungi than for that of bacteria [186].
7.1. Pyrithione
One of the most frequently used zinc ionophore is pyrithione—its antiviral and anticancer properties have already been described in the previous sections. This versatile compound has a long history as a safe and effective compound, which has been used for more than 50 years. It was intensely studied in the 1950s for its fungistatic and bacteriostatic properties. It is one of the ingredients of anti-dandruff shampoos due to its antifungal properties against scalp fungus Malassezia [164].
It is also widely used in other branches of cosmetology because of its broad antimicrobial activity [187]. Nowadays, Zn(II) pyrithione is also known as a compound which neutralizes P. aeruginosa, A. baumannii and S. aureus—bacterial species found in human biofilms, also associated with wound infections resistant to conventional antibiotics. Moreover, zinc pyrithione exhibits a synergistic effect with a leading medicament used to treat wound infections, namely silver sulfadiazine. The combination of these compounds, may effectively inhibit disease progression [188].
7.2. 8-Hydroxyquinoline
Another important zinc ionophore is 8-hydroxyquinoline (PBT2, Figure 4) which often serves as a scaffold or pharmacophore in the design of novel potential drugs with various applications: from antibacterial agents throughout antiviral, antimalarial, antifungal to anticancer and neuroprotecting compounds [189]. It is well known that 8-hydroxyquinoline and its derivatives are excellent metal ion chelators.
They form mono or bidentate complexes, in which heterocyclic nitrogen and phenolate oxygen serve as donor atoms [190,191]. It was shown that these compounds act as a zinc and copper ionophores in mammalian cells [192].
Hydroxyquinoline has been developed as a potential drug for Alzheimer’s and Huntington’s disease, however it failed efficacy checkpoints in the phase 2 human clinical trials [193]. Since PBT2 is an ionophore facilitating the transport of first-row transition metal ions across cell membrane, it alters intracellular metal homeostasis and thus exhibits antimicrobial activity.
The precise mode of its antimicrobial action remains unclear, but involves at least few mechanisms, which makes the development of PBT2 resistance unlikely. The bactericidal mechanisms of its action include: (i) binding of extracellular zinc, transporting metal ion throughout biological membrane and dissociation within the cytoplasm.
A concentration-dependent zinc accumulation with increasing concentrations of PBT2 is observed. Moreover, it was shown that increasing intracellular zinc(II) levels has no effect on the membrane potential, making the whole process electroneutral, (ii) dysregulation of the mtuABC expression and manganese depletion and (iii)disturbance of the intracellular redox balance and induction of oxidative stress via accumulation of reactive oxygen species.
Moreover, antioxidant capacity of the cell is diminished due to decreased level of manganese, making the cell more susceptible to toxic hydrogen peroxide and superoxide anion radical. As a result of Fenton reaction, hydroxyl radical may be formed, leading to oxidative damage of DNA and causing lethal effect to the bacterial cell [194].
7.3. Zincophorin
Many polyoxygenatedionophore-containing natural products exhibit potential antibiotic properties. One of them is zincophorin (Figure 4)—the one whose methyl ester turned out to be an inhibitor of the influenza virus (as mentioned in Section 5).
It is a member of polyketide antibiotic family and possesses high activity against Gram-positive bacteria, including Clostridium perfringens. Antimicrobial activity of zincophorin has been extended to Streptococcus pneumonia lately [195], the leading cause of bacterial pneumonia [196].
In general, minimal inhibitory concentration (MIC) and minimal biofilm inhibitory concentration (MBIC) of zincophorin for three reference strains and four clinical isolates of S. pneumoniae are lower than for daptomycin [196] (a relatively new lipopeptide antibiotic used for the treatment of gram-positive life-threatening infections) [197].
It is worth mentioning that zincophorin losses its antimicrobial activity upon esterification of its carboxylate group. This functional group is crucial both for the biological effect (antibacterial and cytotoxic) and for metal ion binding [189]. Zincophorin is able to bind divalent cations, with the stability order of Zn2+ ≈ Cd2+ > Mg2+ > Sr2+ ≈ Ba2+ ≈ Ca2+ [182].
7.4. Pyrrolidine Dithiocarbamate
As already mentioned, pyrrolidine dithiocarbamate (PDTC) is a versatile compound with antiviral and antibacterial properties [179]. Due to the presence of two thiol groups in its structure, PDTC has heavy metal-chelating and free radical-scavenging properties. Moreover, PDTC is widely used to inhibit the expression of inflammatory genes [198].
While the coexistence of microorganism infection and inflammatory response are the cause of many diseases, finding the compounds having an appropriate dual mode of action is highly desired [199]. Recent studies revealed that PDTC also exhibits antibacterial activity. It inhibits bacterial growth, e.g., that of P. gingivalis (low dose complete response, MIC = 1 mM), A. actinomycetemcomitans and S. aureus (30-times higher dose, MIC = 30 mM).
The antibacterial activity of PDTC is reduced by the addition of copper(II) ions. In contrast, it is enhanced significantly by zinc(II) ions. Obtained results suggest that zinc ions are necessary for antibacterial activity of PDTC, but the precise mechanism of action is not fully elucidated and a more detailed explanation is needed. Nevertheless, PDTC as an anti-inflammatory-antimicrobial-antiviral agent may be useful in the treatment of inflammatory diseases of unknown (bacterial/viral) origin [200].
8. Conclusions
Connecting the fact that Zn(II) is an RNA-dependent RNA polymerase inhibitor and the fact that CQ and HCQ are Zn(II) ionophores leads to an avalanche of concepts. We have comprehensively reviewed the latest available information, describing details of the enzyme itself, showing where Zn(II) and CQ/HCQ (taken together, or independently) show antiviral, and in particular–anticoronaviral action.
FDA–approved Zn(II) ionophores (or even those which are present in food) with antiviral, but also anticancer and antibacterial properties are intensively discussed. The exponentially growing amount of CQ/HCQ-zinc(II) clinical trials is shown within the framework of other ongoing trials.
So, is Zn(II) the éminence grise? The evidence shown in this work let us think that Zn(II) indeed may play a role in the viral cell cycle, although the mechanism still remains uncertain. The idea of zincophore-mediated Zn(II) uptake is definitely one of the well-defined paths that could lead to the development of new anticoronaviral therapeutics.
Although we cannot give a simple one-word answer to the question in the title, we can definitely confirm that it is necessary to continue working on Zn ionophores as potential anti-SARS-CoV-2 agents. It is important to underline once again that this work by no means serves as a piece of medical advice and does not confirm that taking Zn(II) supplements and eating quercetin-rich raspberries/onions/wine will make anyone more immune to SARS-CoV-2.
However, as chemists, we hypothesise that there could be such a possibility and if 24 weeks of clinical trials on CQ/HCQ and Zn(II) [69,70,71] turn out to be a success, this could be a very good direction to go–looking for safer zinc(II) ionophores with less side effects.
Supplementary Materials
An additional table summarizing therapeutics used in anti-COVID-19 clinical trials. This material is available free of charge via the Internet at http://pubs.acs.org. The following are available online at https://www.mdpi.com/1424-8247/13/9/228/s1, Table S1: Summary of non-CQ and non-HCQ based therapeutics in anti-COVID-19 clinical trials.
Author Contributions
Conceptualization, A.H., M.O., K.S.-S., J.W., D.D., A.M., S.P., A.M.-W., A.D.-M., H.K., M.R.-Ż.; formal analysis/data collection, A.H., M.O., K.S.-S., J.W., D.D., A.M., S.P., A.M.-W., A.D.-M., H.K., M.-R.-Ż.; writing—original draft preparation, A.H., M.O., K.S.-S., J.W., D.D., A.M., S.P., A.M.-W., A.D.-M., H.K., M.-R.-Ż.; writing—review and editing, A.H., M.O., K.S.-S., J.W., D.D., A.M., M.-R.-Ż.; visualization, A.H, M.O., K.S.-S., J.W., D.D., A.M.; supervision, M.-R.-Ż.; funding acquisition, H.K., M.-R.-Ż. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support by the National Science Centre (UMO-2017/26/A/ST5/00363 and UMO-2017/26/A/ST5/00364) is gratefully acknowledged.
Acknowledgments
The authors would like to thank Agnieszka Michalczuk for providing us with her artistic vision of SARS-CoV-2.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
| ACE2 | angiotensin-converting enzyme 2; |
| CCHFV | Crimean–Congo hemorrhagic fever virus; |
| CHIKV | Chikungunya virus; |
| DENV | Dengue virus; |
| DTC | dithiocarbamate; |
| EAV | equine arteritis virus; |
| EBOV | Ebola virus; |
| EC50 | effective statistically calculated concentration that induces a specific effect in 50% of experimental population under direct/specific conditions; |
| FCoV | feline coronavirus; |
| FIP | feline infectious peritonitis; |
| FIPV | feline infectious peritonitis virus; |
| FMDV | foot-and-mouth disease virus; |
| HA | hemagglutinin; |
| HAV | hepatitis A; |
| HBsAg | Hepatitis B surface antigen (HBsAg), |
| HBeAg | Hepatitis B antigen; |
| HBV | hepatitis B; |
| HCQ | hydroxychloroquine; |
| HCV | hepatitis C; |
| HIF-1 | hypoxia-inducible factor-1; |
| HIV | human immunodeficiency virus; |
| HK | hinokitol; |
| HSV | herpes simplex virus; |
| IC50 | inhibitory concentration: median value of inhibitor, where 50% of biological and biochemical function of organism inhibition is observed; |
| JEV | Japanese encephalitis virus; |
| NiRAN | nidovirus RdRp-associated nucleotidyltransferase; |
| NTP | nucleoside triphosphate; |
| PDTC | pyrrolidine dithiocarbamate; |
| PRRSV | porcine reproductive and respiratory syndrome virus; |
| PT | pyrithione; |
| RdRp | RNA-dependent RNA polymerase; |
| SARS-CoV | respiratory syndrome coronavirus; |
| SOD | superoxide dismutase-1; |
| UPS | ubiquitin-proteasome system; |
| ZIKV | Zika virus |
References
- Te Velthuis, A.J.W.; van den Worm, S.H.E.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn2+ Inhibits Coronavirus and Arterivirus RNA Polymerase Activity In Vitro and Zinc Ionophores Block the Replication of These Viruses in Cell Culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef]
- Xue, J.; Moyer, A.; Peng, B.; Wu, J.; Hannafon, B.N.; Ding, W.-Q. Chloroquine Is a Zinc Ionophore. PLoS ONE 2014, 9, e109180. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Tian, Z.; Yang, X. Breakthrough: Chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies. Biosci. Trends 2020, 14, 72–73. [Google Scholar] [CrossRef]
- Gautret, P.; Lagier, J.-C.; Parola, P.; van Hoang, T.; Meddeb, L.; Mailhe, M.; Doudier, B.; Courjon, J.; Giordanengo, V.; Vieira, V.E.; et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents 2020, 56, 105949. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Li, C.; Chen, P.; Zhou, N.; Wang, L.; Li, J.; Jiang, H.; Wang, D.-W. Erratum to: Low dose of hydroxychloroquine reduces fatality of critically ill patients with COVID-19. Sci. China Life Sci. 2020. [Google Scholar] [CrossRef]
- Mahévas, M.; Tran, V.-T.; Roumier, M.; Chabrol, A.; Paule, R.; Guillaud, C.; Fois, E.; Lepeule, R.; Szwebel, T.-A.; Lescure, F.-X.; et al. Clinical efficacy of hydroxychloroquine in patients with covid-19 pneumonia who require oxygen: Observational comparative study using routine care data. BMJ 2020, 369, m1844. [Google Scholar] [CrossRef] [PubMed]
- Mehra, M.R.; Desai, S.S.; Ruschitzka, F.; Patel, A.N. RETRACTED: Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: A multinational registry analysis. Lancet 2020. [Google Scholar] [CrossRef]
- World Health Organization. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19—25 May 2020. Available online: https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19—25-may-2020 (accessed on 7 July 2020).
- The Lancet Editors. Expression of concern: Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: A multinational registry analysis. Lancet 2020, 395, e102. [Google Scholar] [CrossRef]
- Mehra, M.R.; Ruschitzka, F.; Patel, A.N. Retraction—Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: A multinational registry analysis. Lancet 2020, 395, 1820. [Google Scholar] [CrossRef]
- Mehra, M.R.; Desai, S.S.; Kuy, S.; Henry, T.D.; Patel, A.N. Cardiovascular Disease, Drug Therapy, and Mortality in Covid-19. N. Engl. J. Med. 2020, 382, e102. [Google Scholar] [CrossRef]
- Scholz, M.; Derwand, R. Does Zinc Supplementation Enhance the Clinical Efficacy of Chloroquine/Hydroxychloroquine to Win Todays Battle Against COVID-19? Preprints 2020, 109815. [Google Scholar] [CrossRef]
- Scholz, M.; Derwand, R.; Zelenko, V. COVID-19 Outpatients–Early Risk–Stratified Treatment with Zinc Plus Low Dose Hydroxychloroquine and Azithromycin: A Retrospective Case Series Study. Preprints 2020, 2020070025. [Google Scholar] [CrossRef]
- Carlucci, P.; Ahuja, T.; Petrilli, C.M.; Rajagopalan, H.; Jones, S.; Rahimian, J. Hydroxychloroquine and azithromycin plus zinc vs. hydroxychloroquine and azithromycin alone: Outcomes in hospitalized COVID-19 patients. medRixv 2020. [Google Scholar] [CrossRef]
- Ahlquist, P.; Noueiry, A.O.; Lee, W.-M.; Kushner, D.B.; Dye, B.T. Host Factors in Positive-Strand RNA Virus Genome Replication. JVI 2003, 77, 8181–8186. [Google Scholar] [CrossRef]
- Miller, W.A.; Koev, G. Synthesis of Subgenomic RNAs by Positive-Strand RNA Viruses. Virology 2000, 273, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Leveque, V.; Ma, H.; Johnson, K.A.; Klumpp, K. Assembly, purification, and pre-steady-state kinetic analysis of active RNA-dependent RNA polymerase elongation complex. J. Biol. Chem. 2012, 287, 10674–10683. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef]
- Stadler, K.; Masignani, V.; Eickmann, M.; Becker, S.; Abrignani, S.; Klenk, H.-D.; Rappuoli, R. SARS–beginning to understand a new virus. Nat. Rev. Microbiol. 2003, 1, 209–218. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest RNA virus genome. Virus Res. 2006, 117, 17–37. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Ward, A.B. Structure of the SARS-CoV nsp12 polymerase bound to nsp7 and nsp8 co-factors. Nat. Commun. 2019, 10, 2342. [Google Scholar] [CrossRef]
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Zhao, Y.; Cao, L.; Wang, T.; Sun, Q.; Ming, Z.; Zhang, L.; et al. Structure of the RNA-dependent RNA polymerase from COVID-19 virus. Science 2020, 368, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, Y.; Weiss, S.; Arnold, E.; Sarafianos, S.G.; Ding, J. Molecular model of SARS coronavirus polymerase: Implications for biochemical functions and drug design. Nucleic Acids Res. 2003, 31, 7117–7130. [Google Scholar] [CrossRef] [PubMed]
- Clercq, E. Antivirals and antiviral strategies. Nat. Rev. Microbiol. 2004, 2, 704–720. [Google Scholar] [CrossRef] [PubMed]
- Uchide, N.; Ohyama, K.; Bessho, T.; Yuan, B.; Yamakawa, T. Effect of antioxidants on apoptosis induced by influenza virus infection: Inhibition of viral gene replication and transcription with pyrrolidine dithiocarbamate. Antivir. Res. 2002, 56, 207–217. [Google Scholar] [CrossRef]
- Suara, R.O.; Crowe, J.E. Effect of zinc salts on respiratory syncytial virus replication. Antimicrob. Agents Chemother. 2004, 48, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Gaudernak, E.; Seipelt, J.; Triendl, A.; Grassauer, A.; Kuechler, E. Antiviral Effects of Pyrrolidine Dithiocarbamate on Human Rhinoviruses. JVI 2002, 76, 6004–6015. [Google Scholar] [CrossRef]
- Si, X.; McManus, B.M.; Zhang, J.; Yuan, J.; Cheung, C.; Esfandiarei, M.; Suarez, A.; Morgan, A.; Luo, H. Pyrrolidine dithiocarbamate reduces coxsackievirus B3 replication through inhibition of the ubiquitin-proteasome pathway. JVI 2005, 79, 8014–8023. [Google Scholar] [CrossRef]
- Korant, B.D.; Kauer, J.C.; Butterworth, B.E. Zinc ions inhibit replication of rhinoviruses. Nature 1974, 248, 588–590. [Google Scholar] [CrossRef]
- Polatnick, J.; Bachrach, H.L. Effect of Zinc and Other Chemical Agents on Foot-and-Mouth Disease Virus Replication. Antimicrob. Agents Chemother. 1978, 13, 731–734. [Google Scholar] [CrossRef]
- Lanke, K.; Krenn, B.M.; Melchers, W.J.G.; Seipelt, J.; van Kuppeveld, F.J.M. PDTC inhibits picornavirus polyprotein processing and RNA replication by transporting zinc ions into cells. J. Gen. Virol. 2007, 88, 1206–1217. [Google Scholar] [CrossRef]
- Krenn, B.M.; Gaudernak, E.; Holzer, B.; Lanke, K.; van Kuppeveld, F.J.M.; Seipelt, J. Antiviral Activity of the Zinc Ionophores Pyrithione and Hinokitiol against Picornavirus Infections. JVI 2009, 83, 58–64. [Google Scholar] [CrossRef]
- Haraguchi, Y.; Sakurai, H.; Hussain, S.; Anner, B.M.; Hoshino, H. Inhibition of HIV-1 infection by zinc group metal compounds. Antivir. Res. 1999, 43, 123–133. [Google Scholar] [CrossRef]
- Katz, E.; Margalith, E. Inhibition of vaccinia virus maturation by zinc chloride. Antimicrob. Agents Chemother. 1981, 19, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Kümel, G.; Schrader, S.; Zentgraf, H.; Daus, H.; Brendel, M. The mechanism of the antiherpetic activity of zinc sulphate. J. Gen. Virol. 1990, 71 Pt 12, 2989–2997. [Google Scholar] [CrossRef]
- Korant, B.D.; Butterworth, B.E. Inhibition by zinc of rhinovirus protein cleavage: Interaction of zinc with capsid polypeptides. JVI 1976, 18, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Novick, S.G.; Godfrey, J.C.; Godfrey, N.J.; Wilder, H.R. How does zinc modify the common cold? Clinical observations and implications regarding mechanisms of action. Med. Hypotheses 1996, 46, 295–302. [Google Scholar] [CrossRef]
- Oxford, J.S.; Perrin, D.D. Inhibition of the particle-associated RNA-dependent RNA polymerase activity of influenza viruses by chelating agents. J. Gen. Virol. 1974, 23, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Wright-Minogue, J.; Fang, J.W.S.; Baroudy, B.M.; Lau, J.Y.N.; Hong, Z. Characterization of Soluble Hepatitis C Virus RNA-Dependent RNA Polymerase Expressed in Escherichia coli. JVI 1999, 73, 1649–1654. [Google Scholar] [CrossRef]
- Hung, M.; Gibbs, C.S.; Tsiang, M. Biochemical characterization of rhinovirus RNA-dependent RNA polymerase. Antivir. Res. 2002, 56, 99–114. [Google Scholar] [CrossRef]
- The PyMOL Molecular Graphics System; Version 1.8; Schrödinger, LLC: New York, NY, USA, 2002.
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Cao, L.; Wang, T.; Wang, Q.; Lou, Z.; Rao, Z. SARS-CoV-2 RNA-Dependent RNA Polymerase in Complex with Cofactors in Reduced Condition. Available online: https://www.rcsb.org/structure/7btf (accessed on 15 April 2020).
- Liu, J.; Cao, R.; Xu, M.; Wang, X.; Zhang, H.; Hu, H.; Li, Y.; Hu, Z.; Zhong, W.; Wang, M. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. 2020, 6, 269. [Google Scholar] [CrossRef]
- McChesney, E.W. Animal toxicity and pharmacokinetics of hydroxychloroquine sulfate. Am. J. Med. 1983, 75, 11–18. [Google Scholar] [CrossRef]
- COVID-19: Reminder of Risk of Serious Side Effects with Chloroquine and Hydroxychloroquine. Available online: http://urpl.gov.pl/sites/default/files/CQ-HCQ%20-%2004-2020%20-%20public%20health%20communication_0.pdf (accessed on 23 April 2020).
- Rainsford, K.D.; Parke, A.L.; Clifford-Rashotte, M.; Kean, W.F. Therapy and pharmacological properties of hydroxychloroquine and chloroquine in treatment of systemic lupus erythematosus, rheumatoid arthritis and related diseases. Inflammopharmacology 2015, 23, 231–269. [Google Scholar] [CrossRef] [PubMed]
- Krafts, K.; Hempelmann, E.; Skórska-Stania, A. From methylene blue to chloroquine: A brief review of the development of an antimalarial therapy. Parasitol. Res. 2012, 111, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mercer, E.; Rekedal, L.; Garg, R.; Lu, B.; Massarotti, E.M.; Solomon, D.H. Hydroxychloroquine improves insulin sensitivity in obese non-diabetic individuals. Arthritis Res. Ther. 2012, 14, R135. [Google Scholar] [CrossRef] [PubMed]
- Solomon, D.H.; Massarotti, E.; Garg, R.; Liu, J.; Canning, C.; Schneeweiss, S. Association between disease-modifying antirheumatic drugs and diabetes risk in patients with rheumatoid arthritis and psoriasis. JAMA 2011, 305, 2525–2531. [Google Scholar] [CrossRef]
- Thomé, R.; Lopes, S.C.P.; Costa, F.T.M.; Verinaud, L. Chloroquine: Modes of action of an undervalued drug. Immunol. Lett. 2013, 153, 50–57. [Google Scholar] [CrossRef]
- Savarino, A.; Boelaert, J.R.; Cassone, A.; Majori, G.; Cauda, R. Effects of chloroquine on viral infections: An old drug against today’s diseases. Lancet Infect. Dis. 2003, 3, 722–727. [Google Scholar] [CrossRef]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef]
- Keyaerts, E.; Vijgen, L.; Maes, P.; Neyts, J.; van Ranst, M. In vitro inhibition of severe acute respiratory syndrome coronavirus by chloroquine. Biochem. Biophys. Res. Commun. 2004, 323, 264–268. [Google Scholar] [CrossRef]
- Zumla, A.; Chan, J.F.W.; Azhar, E.I.; Hui, D.S.C.; Yuen, K.-Y. Coronaviruses-drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef]
- Vincent, M.J.; Bergeron, E.; Benjannet, S.; Erickson, B.R.; Rollin, P.E.; Ksiazek, T.G.; Seidah, N.G.; Nichol, S.T. Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol. J. 2005, 2, 69. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Shen, H.-M. Targeting the Endocytic Pathway and Autophagy Process as a Novel Therapeutic Strategy in COVID-19. Int. J. Biol. Sci. 2020, 16, 1724–1731. [Google Scholar] [CrossRef]
- Millet, J.K.; Whittaker, G.R. Physiological and molecular triggers for SARS-CoV membrane fusion and entry into host cells. Virology 2018, 517, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ (accessed on 15 April 2020).
- de Wilde, A.H.; Jochmans, D.; Posthuma, C.C.; Zevenhoven-Dobbe, J.C.; van Nieuwkoop, S.; Bestebroer, T.M.; van den Hoogen, B.G.; Neyts, J.; Snijder, E.J. Screening of an FDA-Approved Compound Library Identifies Four Small-Molecule Inhibitors of Middle East Respiratory Syndrome Coronavirus Replication in Cell Culture. Antimicrob. Agents Chemother. 2014, 58, 4875–4884. [Google Scholar] [CrossRef]
- Goldman, L.; La Cecil, R.F. Goldman’s Cecil Medicine, 24th ed.; Saunders: Philadelphia, PA, USA, 2011; ISBN 9781437736083. [Google Scholar]
- Pelczar, M.J. Microbiology (An Application Based Approach), 1st ed.; Tata McGraw Hill Education Private Limited: New Delhi, India, 2010; ISBN 978-0-07-015147-5. [Google Scholar]
- Keyaerts, E.; Li, S.; Vijgen, L.; Rysman, E.; Verbeeck, J.; van Ranst, M.; Maes, P. Antiviral Activity of Chloroquine against Human Coronavirus OC43 Infection in Newborn Mice. Antimicrob. Agents Chemother. 2009, 53, 3416–3421. [Google Scholar] [CrossRef]
- Kono, M.; Tatsumi, K.; Imai, A.M.; Saito, K.; Kuriyama, T.; Shirasawa, H. Inhibition of human coronavirus 229E infection in human epithelial lung cells (L132) by chloroquine: Involvement of p38 MAPK and ERK. Antivir. Res. 2008, 77, 150–152. [Google Scholar] [CrossRef]
- Kumar, R.; Khandelwal, N.; Thachamvally, R.; Tripathi, B.N.; Barua, S.; Kashyap, S.K.; Maherchandani, S.; Kumar, N. Role of MAPK/MNK1 signaling in virus replication. Virus Res. 2018, 253, 48–61. [Google Scholar] [CrossRef]
- Hartmann, K. Feline infectious peritonitis. Vet. Clin. N. Am. Small Anim. Pract. 2005, 35, 39–79. [Google Scholar] [CrossRef]
- Takano, T.; Katoh, Y.; Doki, T.; Hohdatsu, T. Effect of chloroquine on feline infectious peritonitis virus infection in vitro and in vivo. Antivir. Res. 2013, 99, 100–107. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. A Study of Hydroxychloroquine, Vitamin C, Vitamin D, and Zinc for the Prevention of COVID-19 Infection (HELPCOVID-19). Available online: https://clinicaltrials.gov/ct2/show/NCT04335084 (accessed on 15 April 2020).
- ClinicalTrials.gov. Proflaxis Using Hydroxychloroquine Plus Vitamins-Zinc During COVID-19 Pandemia. Available online: https://clinicaltrials.gov/ct2/show/NCT04326725 (accessed on 15 April 2020).
- ClinicalTrials.gov. A Study of Quintuple Therapy to Treat COVID-19 Infection (HAZDpaC). Available online: https://clinicaltrials.gov/ct2/show/NCT04334512 (accessed on 15 April 2020).
- ClinicalTrials.gov. Anti-inflammatory/Antioxidant Oral Nutrition Supplementation in COVID-19 (ONSCOVID19). Available online: https://clinicaltrials.gov/ct2/show/NCT04323228 (accessed on 15 April 2020).
- ClinicalTrials.gov. Coronavirus 2019 (COVID-19)-Using Ascorbic Acid and Zinc Supplementation (COVIDAtoZ). Available online: https://clinicaltrials.gov/ct2/show/NCT04342728 (accessed on 15 April 2020).
- ClinicalTrials.gov. Hydroxychloroquine and Zinc with Either Azithromycin or Doxycycline for Treatment of COVID-19 in Outpatient Setting. Available online: https://clinicaltrials.gov/ct2/show/NCT04370782 (accessed on 7 July 2020).
- ClinicalTrials.gov. A Study of Hydroxychloroquine and Zinc in the Prevention of COVID-19 Infection in Military Healthcare Workers (COVID-Milit). Available online: https://clinicaltrials.gov/ct2/show/NCT04377646 (accessed on 7 July 2020).
- ClinicalTrials.gov. Impact of Zinc and Vitamin D3 Supplementation on the Survival of Aged Patients Infected With COVID-19 (ZnD3-CoVici). Available online: https://clinicaltrials.gov/ct2/show/NCT04351490 (accessed on 7 July 2020).
- ClinicalTrials.gov. Early Intervention in COVID-19: Favipiravir Verses Standard Care (PIONEER). Available online: https://clinicaltrials.gov/ct2/show/NCT04373733 (accessed on 28 August 2020).
- ClinicalTrials.gov. Zinc with Chloroquine/Hydroxychloroquine in Treatment of COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04447534 (accessed on 7 July 2020).
- ClinicalTrials.gov. International ALLIANCE Study of Therapies to Prevent Progression of COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04395768 (accessed on 7 July 2020).
- ClinicalTrials.gov. COVID-19 Prophylaxis with Hydroxychloroquine Associated with Zinc for High-Risk Healthcare Workers. Available online: https://clinicaltrials.gov/ct2/show/NCT04384458 (accessed on 7 July 2020).
- ClinicalTrials.gov. A Preventive Treatment for Migrant Workers at High-risk of Covid-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04446104 (accessed on 7 July 2020).
- ClinicalTrials.gov. New Antiviral Drugs for Treatment of COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04392427 (accessed on 7 July 2020).
- ClinicalTrials.gov. Covid-19 and Diabetes in West of Algeria (COVIDIAB-13). Available online: https://clinicaltrials.gov/ct2/show/NCT04412746 (accessed on 7 July 2020).
- ClinicalTrials.gov. The Study of Quadruple Therapy Zinc, Quercetin, Bromelain and Vitamin C on the Clinical Outcomes of Patients Infected with COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04468139 (accessed on 30 July 2020).
- ClinicalTrials.gov. Efficacy of Subcutaneous Ivermectin with or without Zinc and Nigella Sativa in COVID-19 Patients (SINZ-COVID-PK). Available online: https://clinicaltrials.gov/ct2/show/NCT04472585 (accessed on 30 July 2020).
- ClinicalTrials.gov. Clearing the Fog: Is Hydroxychloroquine Effective in Reducing COVID-19 Progression (COVID-19). Available online: https://clinicaltrials.gov/ct2/show/NCT04491994 (accessed on 30 July 2020).
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Milewska, A.; Chi, Y.; Szczepanski, A.; Barreto-Duran, E.; Liu, K.; Liu, D.; Guo, X.; Ge, Y.; Li, J.; Cui, L.; et al. HTCC as a highly effective polymeric inhibitor of SARS-CoV-2 and MERS-CoV. bioRxiv 2020. [Google Scholar] [CrossRef]
- Rut, W.; Groborz, K.; Zhang, L.; Sun, X.; Zmudzinski, M.; Hilgenfeld, R.; Drag, M. Substrate specificity profiling of SARS-CoV-2 M pro protease provides basis for anti-COVID-19 drug design. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef]
- Han, Y.-S.; Chang, G.-G.; Juo, C.-G.; Lee, H.-J.; Yeh, S.-H.; Hsu, J.T.-A.; Chen, X. Papain-like protease 2 (PLP2) from severe acute respiratory syndrome coronavirus (SARS-CoV): Expression, purification, characterization, and inhibition. Biochemistry 2005, 44, 10349–10359. [Google Scholar] [CrossRef]
- Imbert, I.; Guillemot, J.-C.; Bourhis, J.-M.; Bussetta, C.; Coutard, B.; Egloff, M.-P.; Ferron, F.; Gorbalenya, A.E.; Canard, B. A second, non-canonical RNA-dependent RNA polymerase in SARS coronavirus. EMBO J. 2006, 25, 4933–4942. [Google Scholar] [CrossRef]
- Wu, Y.-S.; Lin, W.-H.; Hsu, J.T.-A.; Hsieh, H.-P. Antiviral drug discovery against SARS-CoV. Curr. Med. Chem. 2006, 13, 2003–2020. [Google Scholar] [CrossRef]
- Adedeji, A.O.; Marchand, B.; te Velthuis, A.J.W.; Snijder, E.J.; Weiss, S.; Eoff, R.L.; Singh, K.; Sarafianos, S.G. Mechanism of nucleic acid unwinding by SARS-CoV helicase. PLoS ONE 2012, 7, e36521. [Google Scholar] [CrossRef]
- Boelaert, J.R.; Piette, J.; Sperber, K. The potential place of chloroquine in the treatment of HIV-1-infected patients. J. Clin. Virol. 2001, 20, 137–140. [Google Scholar] [CrossRef]
- Savarino, A.; Gennero, L.; Sperber, K.; Boelaert, J.R. The anti-HIV-1 activity of chloroquine. J. Clin. Virol. 2001, 20, 131–135. [Google Scholar] [CrossRef]
- Naarding, M.A.; Baan, E.; Pollakis, G.; Paxton, W.A. Effect of chloroquine on reducing HIV-1 replication in vitro and the DC-SIGN mediated transfer of virus to CD4 + T-lymphocytes. Retrovirology 2007, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Savarino, A.; Lucia, M.B.; Rastrelli, E.; Rutella, S.; Golotta, C.; Morra, E.; Tamburrini, E.; Perno, C.F.; Boelaert, J.R.; Sperber, K.; et al. Anti-HIV Effects of Chloroquine: Inhibition of viral particle glycosylation and synergism with protease inhibitors. J. Acquir. Immune Defic. Syndr. 2004, 35, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, F.; Smith, K.; Hoven, A. Chloroquine and Hydroxychloroquine as Inhibitors of Human Immunodeficiency Virus (HIV-1) Activity. Curr. Pharm. Des. 2004, 10, 2643–2648. [Google Scholar] [CrossRef]
- Savarino, A.; Gennero, L.; Chen, H.C.; Serrano, D.; Malavasi, F.; Boelaert, J.R.; Sperber, K. Anti-HIV effects of chloroquine: Mechanisms of inhibition and spectrum of activity. J. Acquir. Immune Defic. Syndr. 2001, 15, 2221–2229. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Yang, J.; Diagne, A. Chloroquine inhibits HIV-1 replication in human peripheral blood lymphocytes. Immunol. Lett. 1998, 64, 45–47. [Google Scholar] [CrossRef]
- Bishop, N.E. Examination of potential inhibitors of hepatitis A virus uncoating. Intervirology 1998, 41, 261–271. [Google Scholar] [CrossRef]
- Kouroumalis, E.A.; Koskinas, J. Treatment of chronic active hepatitis B (CAH B) with chloroquine: A preliminary report. Ann. Acad. Med. Singap. 1986, 15, 149–152. [Google Scholar]
- Mizui, T.; Yamashina, S.; Tanida, I.; Takei, Y.; Ueno, T.; Sakamoto, N.; Ikejima, K.; Kitamura, T.; Enomoto, N.; Sakai, T.; et al. Inhibition of hepatitis C virus replication by chloroquine targeting virus-associated autophagy. J. Gastroenterol. 2010, 45, 195–203. [Google Scholar] [CrossRef]
- Moraes Falcão, L.; Terrabuio, D.R.B.; Diniz, M.A.; da Silva Evangelista, A.; Souza, F.G.; R Cancado, E.L. Efficacy and safety of chloroquine plus prednisone for the treatment of autoimmune hepatitis in a randomized trial. JGH Open 2020, 4, 371–377. [Google Scholar] [CrossRef]
- Kronenberger, P.; Vrijsen, R.; Boeyé, A. Chloroquine induces empty capsid formation during poliovirus eclipse. JVI 1991, 65, 7008–7011. [Google Scholar] [CrossRef] [PubMed]
- Ooi, E.E.; Chew, J.S.W.; Loh, J.P.; Chua, R.C.S. In vitro inhibition of human influenza a virus replication by chloroquine. Virol. J. 2006, 3, 39. [Google Scholar] [CrossRef] [PubMed]
- Vigerust, D.J.; McCullers, J.A. Chloroquine is effective against influenza A virus in vitro but not in vivo. Influenza Other Respir. Viruses 2007, 1, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Aoki, H.; Tsurumi, T.; Sugiura, Y.; Nishiyama, Y.; Suzuki, S.; Maeno, K. Mechanism of Uncoating of Influenza B Virus in MDCK Cells: Action of Chloroquine. J. Gen. Virol. 1983, 64, 1149–1156. [Google Scholar] [CrossRef]
- Yan, Y.; Zou, Z.; Sun, Y.; Li, X.; Xu, K.-F.; Wei, Y.; Jin, N.; Jiang, C. Anti-malaria drug chloroquine is highly effective in treating avian influenza A H5N1 virus infection in an animal model. Cell Res. 2013, 23, 300–302. [Google Scholar] [CrossRef]
- Delvecchio, R.; Higa, L.M.; Pezzuto, P.; Valadão, A.L.; Garcez, P.P.; Monteiro, F.L.; Loiola, E.C.; Dias, A.A.; Silva, F.J.M.; Aliota, M.T.; et al. Chloroquine, an Endocytosis Blocking Agent, Inhibits Zika Virus Infection in Different Cell Models. Viruses 2016, 8, 322. [Google Scholar] [CrossRef]
- Li, C.; Zhu, X.; Ji, X.; Quanquin, N.; Deng, Y.-Q.; Tian, M.; Aliyari, R.; Zuo, X.; Yuan, L.; Afridi, S.K.; et al. Chloroquine, a FDA-approved Drug, Prevents Zika Virus Infection and its Associated Congenital Microcephaly in Mice. EBioMedicine 2017, 24, 189–194. [Google Scholar] [CrossRef]
- Zhang, S.; Yi, C.; Li, C.; Zhang, F.; Peng, J.; Wang, Q.; Liu, X.; Ye, X.; Li, P.; Wu, M.; et al. Chloroquine inhibits endosomal viral RNA release and autophagy-dependent viral replication and effectively prevents maternal to fetal transmission of Zika virus. Antivir. Res. 2019, 169, 104547. [Google Scholar] [CrossRef]
- Dowall, S.D.; Bosworth, A.; Watson, R.; Bewley, K.; Taylor, I.; Rayner, E.; Hunter, L.; Pearson, G.; Easterbrook, L.; Pitman, J.; et al. Chloroquine inhibited Ebola virus replication in vitro but failed to protect against infection and disease in the in vivo guinea pig model. J. Gen. Virol. 2015, 96, 3484–3492. [Google Scholar] [CrossRef]
- Ferraris, O.; Moroso, M.; Pernet, O.; Emonet, S.; Ferrier Rembert, A.; Paranhos-Baccalà, G.; Peyrefitte, C.N. Evaluation of Crimean-Congo hemorrhagic fever virus in vitro inhibition by chloroquine and chlorpromazine, two FDA approved molecules. Antivir. Res. 2015, 118, 75–81. [Google Scholar] [CrossRef]
- Tsiang, H.; Superti, F. Ammonium chloride and chloroquine inhibit rabies virus infection in neuroblastoma cells. Brief report. Arch. Virol. 1984, 81, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Santhosh, S.R.; Tiwari, M.; Lakshmana Rao, P.V.; Parida, M. Assessment of in vitro prophylactic and therapeutic efficacy of chloroquine against Chikungunya virus in vero cells. J. Med. Virol. 2010, 82, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Delogu, I.; de Lamballerie, X. Chikungunya disease and chloroquine treatment. J. Med. Virol. 2011, 83, 1058–1059. [Google Scholar] [CrossRef] [PubMed]
- Farias, K.J.S.; Machado, P.R.L.; da Fonseca, B.A.L. Chloroquine Inhibits Dengue Virus Type 2 Replication in Vero Cells but Not in C6/36 Cells. Sci. World J. 2013, 2013, 1–5. [Google Scholar] [CrossRef]
- Farias, K.J.S.; Machado, P.R.L.; de Almeida Junior, R.F.; de Aquino, A.A.; da Fonseca, B.A.L. Chloroquine interferes with dengue-2 virus replication in U937 cells. Microbiol. Immunol. 2014, 58, 318–326. [Google Scholar] [CrossRef]
- Farias, K.J.S.; Machado, P.R.L.; Muniz, J.A.P.C.; Imbeloni, A.A.; da Fonseca, B.A.L. Antiviral Activity of Chloroquine Against Dengue Virus Type 2 Replication in Aotus Monkeys. Viral Immunol. 2015, 28, 161–169. [Google Scholar] [CrossRef]
- Freiberg, A.N.; Worthy, M.N.; Lee, B.; Holbrook, M.R. Combined chloroquine and ribavirin treatment does not prevent death in a hamster model of Nipah and Hendra virus infection. Arch. Virol. 2010, 91, 765–772. [Google Scholar] [CrossRef]
- Koyama, A.H.; Uchida, T. Inhibition of multiplication of herpes simplex virus type 1 by ammonium chloride and chloroquine. Virology 1984, 138, 332–335. [Google Scholar] [CrossRef]
- Sperber, K.; Louie, M.; Kraus, T.; Proner, J.; Sapira, E.; Lin, S.; Stecher, V.; Mayer, L. Hydroxychloroquine treatment of patients with human immunodeficiency virus type 1. Clin. Ther. 1995, 17, 622–636. [Google Scholar] [CrossRef]
- Sperber, K.; Chiang, G.; Chen, H.; Ross, W.; Chusid, E.; Gonchar, M.; Chow, R.; Liriano, O. Comparison of hydroxychloroquine with zidovudine in asymptomatic patients infected with human immunodeficiency virus type 1. Clin. Ther. 1997, 19, 913–923. [Google Scholar] [CrossRef]
- Sperber, K.; Kalb, T.H.; Stecher, V.J.; Banerjee, R.; Mayer, L. Inhibition of human immunodeficiency virus type 1 replication by hydroxychloroquine in T cells and monocytes. AIDS Res. Hum. Retrovir. 1993, 9, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Fesen, M.R.; Kohn, K.W.; Leteurtre, F.; Pommier, Y. Inhibitors of human immunodeficiency virus integrase. Proc. Natl. Acad. Sci. USA 1993, 90, 2399–2403. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.C.; Lin, J.K.; Chen, S.S. Inhibition of HIV-1 Tat-mediated transactivation by quinacrine and chloroquine. Biochem. Biophys. Res. Commun. 1996, 226, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Morra, E.; Savarino, A.; Gennero, L.; Pescarmona, G.P. Effects of chloroquine on iron metabolism in a lymphocytic cell line. J. Clin. Virol. 2000, 16, 91–92. [Google Scholar]
- Tsai, W.P.; Nara, P.L.; Kung, H.F.; Oroszlan, S. Inhibition of human immunodeficiency virus infectivity by chloroquine. AIDS Res. Hum. Retrovir. 1990, 6, 481–489. [Google Scholar] [CrossRef]
- Tricou, V.; Minh, N.N.; Van, T.P.; Lee, S.J.; Farrar, J.; Wills, B.; Tran, H.T.; Simmons, C.P. A randomized controlled trial of chloroquine for the treatment of dengue in Vietnamese adults. PLoS Negl. Trop. Dis. 2010, 4, e785. [Google Scholar] [CrossRef]
- Borges, M.C.; Castro, L.A.; Fonseca, B.A.L. Chloroquine use improves dengue-related symptoms. Mem. Inst. Oswaldo Cruz 2013, 108, 596–599. [Google Scholar] [CrossRef]
- Madrid, P.B.; Chopra, S.; Manger, I.D.; Gilfillan, L.; Keepers, T.R.; Shurtleff, A.C.; Green, C.E.; Iyer, L.V.; Dilks, H.H.; Davey, R.A.; et al. A systematic screen of FDA-approved drugs for inhibitors of biological threat agents. PLoS ONE 2013, 8, e60579. [Google Scholar] [CrossRef]
- Guo, C.; Zhu, Z.; Wang, X.; Chen, Y.; Liu, X. Pyrithione inhibits porcine reproductive and respiratory syndrome virus replication through interfering with NF-κB and heparanase. Vet. Microbiol. 2017, 201, 231–239. [Google Scholar] [CrossRef]
- Qiu, M.; Chen, Y.; Chu, Y.; Song, S.; Yang, N.; Gao, J.; Wu, Z. Zinc ionophores pyrithione inhibits herpes simplex virus replication through interfering with proteasome function and NF-κB activation. Antivir. Res. 2013, 100, 44–53. [Google Scholar] [CrossRef]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef] [PubMed]
- Uchide, N.; Toyoda, H. Antioxidant therapy as a potential approach to severe influenza-associated complications. Molecules 2011, 16, 2032–2052. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, R.; Li, X.; He, J.; Jiang, S.; Liu, S.; Yang, J. Quercetin as an Antiviral Agent Inhibits Influenza A Virus (IAV) Entry. Viruses 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Sun, G.; Guo, W.; Huang, Y.; Sun, W.; Zhao, F.; Hu, K. Inhibition of hepatitis B virus replication by quercetin in human hepatoma cell lines. Virol. Sin. 2015, 30, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Rojas, Á.; Del Campo, J.A.; Clement, S.; Lemasson, M.; García-Valdecasas, M.; Gil-Gómez, A.; Ranchal, I.; Bartosch, B.; Bautista, J.D.; Rosenberg, A.R.; et al. Effect of Quercetin on Hepatitis C Virus Life Cycle: From Viral to Host Targets. Sci. Rep. 2016, 6, 55. [Google Scholar] [CrossRef]
- Bachmetov, L.; Gal-Tanamy, M.; Shapira, A.; Vorobeychik, M.; Giterman-Galam, T.; Sathiyamoorthy, P.; Golan-Goldhirsh, A.; Benhar, I.; Tur-Kaspa, R.; Zemel, R. Suppression of hepatitis C virus by the flavonoid quercetin is mediated by inhibition of NS3 protease activity. J. Viral. Hepat. 2012, 19, e81–e88. [Google Scholar] [CrossRef]
- Qiu, X.; Kroeker, A.; He, S.; Kozak, R.; Audet, J.; Mbikay, M.; Chrétien, M. Prophylactic Efficacy of Quercetin 3-β- O -d-Glucoside against Ebola Virus Infection. Antimicrob. Agents Chemother. 2016, 60, 5182–5188. [Google Scholar] [CrossRef]
- Chiow, K.H.; Phoon, M.C.; Putti, T.; Tan, B.K.H.; Chow, V.T. Evaluation of antiviral activities of Houttuynia cordata Thunb. extract, quercetin, quercetrin and cinanserin on murine coronavirus and dengue virus infection. Asian Pac. J. Trop. Med. 2016, 9, 1–7. [Google Scholar] [CrossRef]
- Johari, J.; Kianmehr, A.; Mustafa, M.; Abubakar, S.; Zandi, K. Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef]
- Wong, G.; He, S.; Siragam, V.; Bi, Y.; Mbikay, M.; Chretien, M.; Qiu, X. Antiviral activity of quercetin-3-β-O-D-glucoside against Zika virus infection. Virol. Sin. 2017, 32, 545–547. [Google Scholar] [CrossRef]
- Huang, Q.; Sivaramakrishna, R.P.; Ludwig, K.; Korte, T.; Böttcher, C.; Herrmann, A. Early steps of the conformational change of influenza virus hemagglutinin to a fusion active state. Biochim. Biophys. Acta Biomembr. 2003, 1614, 3–13. [Google Scholar] [CrossRef]
- Gaspari, P.; Banerjee, T.; Malachowski, W.P.; Muller, A.J.; Prendergast, G.C.; DuHadaway, J.; Bennett, S.; Donovan, A.M. Structure-activity study of brassinin derivatives as indoleamine 2,3-dioxygenase inhibitors. J. Med. Chem. 2006, 49, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.-L.; Feng, Y.-P.; Jiang, Y.-Y.; Liu, S.-Y.; Ding, G.-Y.; Li, R.-T. Synthesis and in vitro antitumor activity of 4(3H)-quinazolinone derivatives with dithiocarbamate side chains. Bioorg. Med. Chem. Lett. 2005, 15, 1915–1917. [Google Scholar] [CrossRef]
- Li, Y.; Qi, H.; Li, X.; Hou, X.; Lu, X.; Xiao, X. A novel dithiocarbamate derivative induces cell apoptosis through p53-dependent intrinsic pathway and suppresses the expression of the E6 oncogene of human papillomavirus 18 in HeLa cells. Apoptosis 2015, 20, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, E.C.; Kern, P.; Dietrich, M.; Ernst, M.; Flad, H.D.; Bock, P.; Group, G.D.S. Inhibition of HIV progression by dithiocarb. Lancet 1990, 335, 679–682. [Google Scholar] [CrossRef]
- Brooks, H.A.; Gardner, D.; Poyser, J.P.; King, T.J. The structure and absolute stereochemistry of zincophorin (antibiotic M144255): A monobasic carboxylic acid ionophore having a remarkable specificity for divalent cations. J. Antibiot. 1984, 37, 1501–1504. [Google Scholar] [CrossRef]
- Gräfe, U.; Schade, W.; Roth, M.; Radics, L.; Incze, M.; Ujszászy, K. Griseochelin, a novel carboxylic acid antibiotic from Streptomyces griseus. J. Antibiot. 1984, 37, 836–846. [Google Scholar] [CrossRef]
- Radics, L. The constitution of griseochelin. J. Chem. Soc. Chem. Commun. 1984, 1010, 599. [Google Scholar] [CrossRef]
- Gräfe, U.; Tonew, E.; Schade, W.; Reinhardt, G.; Hartl, A. Ger. (East). Patent DD 231 793 1986. [Google Scholar]
- Tonew, E.; Tonew, M.; Graefe, U.; Zöpel, P. Griseochelinmethylester, ein neues Polyether-Derivat mit antiviraler Wirksamkeit. Pharmazie 1988, 43, 717–719. [Google Scholar]
- Huang, R.; Wallqvist, A.; Covell, D.G. Anticancer metal compounds in NCI’s tumor-screening database: Putative mode of action. Biochem. Pharmacol. 2005, 69, 1009–1039. [Google Scholar] [CrossRef]
- Potocki, S.; Rowinska-Zyrek, M.; Witkowska, D.; Pyrkosz, M.; Szebesczyk, A.; Krzywoszynska, K.; Kozlowski, H. Metal transport and homeostasis within the human body: Toxicity associated with transport abnormalities. Curr. Med. Chem. 2012, 19, 2738–2759. [Google Scholar] [CrossRef] [PubMed]
- Magda, D.; Lecane, P.; Wang, Z.; Hu, W.; Thiemann, P.; Ma, X.; Dranchak, P.K.; Wang, X.; Lynch, V.; Wei, W.; et al. Synthesis and Anticancer Properties of Water-Soluble Zinc Ionophores. Cancer Res. 2008, 68, 5318–5325. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.-Q.; Lind, S.E. Metal ionophores: An emerging class of anticancer drugs. IUBMB Life 2009, 61, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, D.B.; Richardson, D.R. Iron chelators as anti-neoplastic agents: Current developments and promise of the PIH class of chelators. Curr. Med. Chem. 2003, 10, 1035–1049. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J.; Efstathiou, A.; Ioannou-Loucaides, S.; Kolnagou, A. Chelators Controlling Metal Metabolism and Toxicity Pathways: Applications in Cancer Prevention, Diagnosis and Treatment. Hemoglobin 2008, 32, 217–227. [Google Scholar] [CrossRef]
- Gaur, K.; Vázquez-Salgado, A.; Duran-Camacho, G.; Dominguez-Martinez, I.; Benjamín-Rivera, J.; Fernández-Vega, L.; Carmona Sarabia, L.; Cruz García, A.; Pérez-Deliz, F.; Méndez Román, J.; et al. Iron and Copper Intracellular Chelation as an Anticancer Drug Strategy. Inorganics 2018, 6, 126. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, J.H.; Moon, S.J.; Chung, K.C.; Hsu, C.Y.; Seo, J.T.; Ahn, Y.S. Pyrithione, a zinc ionophore, inhibits NF-kappaB activation. Biochem. Biophys. Res. Commun. 1999, 259, 505–509. [Google Scholar] [CrossRef]
- Knight, D.W.; Hartung, J. 1-Hydroxypyridine-2(1H)-thione. In Encyclopedia of Reagents for Organic Synthesis; John Wiley & Sons, Ltd.: Chichester, UK, 2001; p. 1875. ISBN 0471936235. [Google Scholar]
- Feng, P.; Li, T.L.; Guan, Z.X.; Franklin, R.B.; Costello, L.C. Effect of zinc on prostatic tumorigenicity in nude mice. Ann. N. Y. Acad. Sci. 2003, 1010, 316–320. [Google Scholar] [CrossRef]
- Ding, W.-Q.; Liu, B.; Vaught, J.L.; Yamauchi, H.; Lind, S.E. Anticancer activity of the antibiotic clioquinol. Cancer Res. 2005, 65, 3389–3395. [Google Scholar] [CrossRef]
- Ritchie, C.W.; Bush, A.I.; Mackinnon, A.; Macfarlane, S.; Mastwyk, M.; MacGregor, L.; Kiers, L.; Cherny, R.; Li, Q.-X.; Tammer, A.; et al. Metal-protein attenuation with iodochlorhydroxyquin (clioquinol) targeting Abeta amyloid deposition and toxicity in Alzheimer disease: A pilot phase 2 clinical trial. Arch. Neurol. 2003, 60, 1685–1691. [Google Scholar] [CrossRef]
- Misra, H.P. Reaction of copper-zinc superoxide dismutase with diethyldithiocarbamate. J. Biol. Chem. 1979, 254, 11623–11628. [Google Scholar] [PubMed]
- Schreck, R.; Meier, B.; Männel, D.N.; Dröge, W.; Baeuerle, P.A. Dithiocarbamates as potent inhibitors of nuclear factor kappa B activation in intact cells. J. Exp. Med. 1992, 175, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Verhaegh, G.W.; Richard, M.J.; Hainaut, P. Regulation of p53 by metal ions and by antioxidants: Dithiocarbamate down-regulates p53 DNA-binding activity by increasing the intracellular level of copper. Mol. Cell. Biol. 1997, 17, 5699–5706. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.H.; Momand, J. Pyrrolidine dithiocarbamate prevents p53 activation and promotes p53 cysteine residue oxidation. J. Biol. Chem. 1998, 273, 18898–18905. [Google Scholar] [CrossRef] [PubMed]
- Nobel, C.I.; Kimland, M.; Lind, B.; Orrenius, S.; Slater, A.F. Dithiocarbamates induce apoptosis in thymocytes by raising the intracellular level of redox-active copper. J. Biol. Chem. 1995, 270, 26202–26208. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, J.H.; Xu, J.; Hsu, C.Y.; Ahn, Y.S. Pyrrolidine Dithiocarbamate Induces Bovine Cerebral Endothelial Cell Death by Increasing the Intracellular Zinc Level. J. Neurochem. 1999, 72, 1586–1592. [Google Scholar] [CrossRef]
- Puschner, B.; Dechant, J.E. Common Toxins in Equine Practice. In Equine Medicine, 7th ed.; Robinson, N.E., Sprayberry, K.A., Eds.; Elsevier: St. Louis, MO, USA, 2015; pp. 922–927. ISBN 9781455745555. [Google Scholar]
- Patel, M.B.; Garrad, E.; Meisel, J.W.; Negin, S.; Gokel, M.R.; Gokel, G.W. Synthetic ionophores as non-resistant antibiotic adjuvants. RSC Adv. 2019, 9, 2217–2230. [Google Scholar] [CrossRef]
- Rutkowski, J.; Brzezinski, B. Structures and Properties of Naturally Occurring Polyether Antibiotics. Biomed Res. Int. 2013, 2013, 1–31. [Google Scholar] [CrossRef]
- Shen, J.; Brodbelt, J.S. Characterization of ionophore–metal complexes by infrared multiphoton photodissociation and collision activated dissociation in a quadrupole ion trap mass spectrometer. Analyst 2000, 125, 641–650. [Google Scholar] [CrossRef]
- Sugarman, B. Zinc and infection. Rev. Infect. Dis. 1983, 5, 137–147. [Google Scholar] [CrossRef]
- Wątły, J.; Potocki, S.; Rowińska-Żyrek, M. Zinc Homeostasis at the Bacteria/Host Interface-From Coordination Chemistry to Nutritional Immunity. Chemistry 2016, 22, 15992–16010. [Google Scholar] [CrossRef] [PubMed]
- Pasquet, J.; Chevalier, Y.; Pelletier, J.; Couval, E.; Bouvier, D.; Bolzinger, M.-A. The contribution of zinc ions to the antimicrobial activity of zinc oxide. Colloids Surf. A Physicochem. Eng. Asp. 2014, 457, 263–274. [Google Scholar] [CrossRef]
- Schwartz, J.R. Zinc Pyrithione: A Topical Antimicrobial with Complex Pharmaceutics. J. Drugs Dermatol. 2016, 15, 140–144. [Google Scholar] [PubMed]
- Blanchard, C.; Brooks, L.; Ebsworth-Mojica, K.; Didione, L.; Wucher, B.; Dewhurst, S.; Krysan, D.; Dunman, P.M.; Wozniak, R.A.F. Zinc Pyrithione Improves the Antibacterial Activity of Silver Sulfadiazine Ointment. mSphere 2016, 1, 322. [Google Scholar] [CrossRef] [PubMed]
- McGowan, J.E.; Harper, A.D.; Davison, E.K.; Jeong, J.Y.; Mros, S.; Harbison-Price, N.; van Zuylen, E.M.; Knottenbelt, M.K.; Heikal, A.; Ferguson, S.A.; et al. Substituted sulfonamide bioisosteres of 8-hydroxyquinoline as zinc-dependent antibacterial compounds. Bioorg. Med. Chem. Lett. 2020, 30, 127110. [Google Scholar] [CrossRef] [PubMed]
- Di Vaira, M.; Bazzicalupi, C.; Orioli, P.; Messori, L.; Bruni, B.; Zatta, P. Clioquinol, a drug for Alzheimer’s disease specifically interfering with brain metal metabolism: Structural characterization of its zinc(II) and copper(II) complexes. Inorg. Chem. 2004, 43, 3795–3797. [Google Scholar] [CrossRef]
- Alfonso, I.; Quesada, R. Biological activity of synthetic ionophores: Ion transporters as prospective drugs? Chem. Sci. 2013, 4, 3009. [Google Scholar] [CrossRef]
- Yu, H.; Zhou, Y.; Lind, S.E.; Ding, W.-Q. Clioquinol targets zinc to lysosomes in human cancer cells. Biochem. J. 2009, 417, 133–139. [Google Scholar] [CrossRef]
- Mao, X.; Li, X.; Sprangers, R.; Wang, X.; Venugopal, A.; Wood, T.; Zhang, Y.; Kuntz, D.A.; Coe, E.; Trudel, S.; et al. Clioquinol inhibits the proteasome and displays preclinical activity in leukemia and myeloma. Leukemia 2009, 23, 585–590. [Google Scholar] [CrossRef]
- Ding, W.-Q.; Yu, H.-J.; Lind, S.E. Zinc-binding compounds induce cancer cell death via distinct modes of action. Cancer Lett. 2008, 271, 251–259. [Google Scholar] [CrossRef]
- Kavlock, R.J. Pesticides as Endocrine-Disrupting Chemicals. In Methods of Tissue Engineering; Atala, A., Lanza, R.P., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 727–746. ISBN 9780124262607. [Google Scholar]
- Bessho, R.; Matsubara, K.; Kubota, M.; Kuwakado, K.; Hirota, H.; Wakazono, Y.; Lin, Y.W.; Okuda, A.; Kawai, M.; Nishikomori, R.; et al. Pyrrolidine dithiocarbamate, a potent inhibitor of nuclear factor κB (NF-κB) activation, prevents apoptosis in human promyelocytic leukemia HL-60 cells and thymocytes. Biochem. Pharmacol. 1994, 48, 1883–1889. [Google Scholar] [CrossRef]
- Nguyen, M.; Vendier, L.; Stigliani, J.-L.; Meunier, B.; Robert, A. Structures of the Copper and Zinc Complexes of PBT2, a Chelating Agent Evaluated as Potential Drug for Neurodegenerative Diseases. Eur. J. Inorg. Chem. 2017, 2017, 600–608. [Google Scholar] [CrossRef]
- Adlard, P.A.; Cherny, R.A.; Finkelstein, D.I.; Gautier, E.; Robb, E.; Cortes, M.; Volitakis, I.; Liu, X.; Smith, J.P.; Perez, K.; et al. Rapid Restoration of Cognition in Alzheimer’s Transgenic Mice with 8-Hydroxy Quinoline Analogs Is Associated with Decreased Interstitial Aβ. Neuron 2008, 59, 43–55. [Google Scholar] [CrossRef]
- Lannfelt, L.; Blennow, K.; Zetterberg, H.; Batsman, S.; Ames, D.; Harrison, J.; Masters, C.L.; Targum, S.; Bush, A.I.; Murdoch, R.; et al. Safety, efficacy, and biomarker findings of PBT2 in targeting Aβ as a modifying therapy for Alzheimer’s disease: A phase IIa, double-blind, randomised, placebo-controlled trial. Lancet Neurol. 2008, 7, 779–786. [Google Scholar] [CrossRef]
- Harbison-Price, N.; Ferguson, S.A.; Heikal, A.; Taiaroa, G.; Hards, K.; Nakatani, Y.; Rennison, D.; Brimble, M.A.; El-Deeb, I.M.; Bohlmann, L.; et al. Multiple Bactericidal Mechanisms of the Zinc Ionophore PBT2. mSphere 2020, 5, 5649. [Google Scholar] [CrossRef] [PubMed]
- Walther, E.; Boldt, S.; Kage, H.; Lauterbach, T.; Martin, K.; Roth, M.; Hertweck, C.; Sauerbrei, A.; Schmidtke, M.; Nett, M. Zincophorin–biosynthesis in Streptomyces griseus and antibiotic properties. GMS Infect. Dis. 2016, 4, 1–9. [Google Scholar] [CrossRef]
- Farver, C.F.; Zander, D.S. Molecular Basis of Pulmonary Disease. Mol. Pathol. 2009, 417, 305–364. [Google Scholar] [CrossRef]
- Schoen, C.; Unzicker, C.; Stuhler, G.; Elias, J.; Einsele, H.; Grigoleit, G.U.; Abele-Horn, M.; Mielke, S. Life-Threatening Infection Caused by Daptomycin-Resistant Corynebacterium jeikeium in a Neutropenic Patient. J. Clin. Microbiol. 2009, 47, 2328–2331. [Google Scholar] [CrossRef]
- Mochizuki, T.; Satsu, H.; Shimizu, M. Signaling pathways involved in tumor necrosis factor alpha-induced upregulation of the taurine transporter in Caco-2 cells. FEBS Lett. 2005, 579, 3069–3074. [Google Scholar] [CrossRef]
- Sharma, P.K.; Kumar, S.; Kumar, P.; Kaushik, P.; Kaushik, D.; Dhingra, Y.; Aneja, K.R. Synthesis and biological evaluation of some pyrazolylpyrazolines as anti-inflammatory–antimicrobial agents. Eur. J. Med. Chem. 2010, 45, 2650–2655. [Google Scholar] [CrossRef]
- Kang, M.-S.; Choi, E.-K.; Choi, D.-H.; Ryu, S.-Y.; Lee, H.-H.; Kang, H.-C.; Koh, J.-T.; Kim, O.-S.; Hwang, Y.-C.; Yoon, S.-J.; et al. Antibacterial activity of pyrrolidine dithiocarbamate. FEMS Microbiol. Lett. 2008, 280, 250–254. [Google Scholar] [CrossRef]
Related:
Hydroxychloroquine and azithromycin tolerance in haemodialysis patients during COVID-19 infection